На групповой состав пирофосфатных экстрактов
(по Кононовой-Бельчиковой)
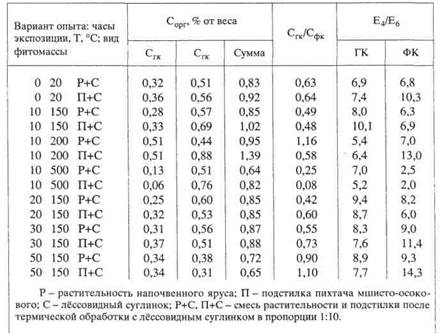
ном эксперименте с насыпным профилем из материала реальных горизонтов подзолистой почвы в стеклянной колонке (рис. 7). На поверхности "почвы" из реальных горизонтов поместили послойно снизу вверх естественную, обугленную и озоленную подстилку пихтача мшисто-осокового. Для возбуждения микробиологической активности ввели, по совету микробиолога И.Л. Клевен-ской, питательную смесь (5% раствор глюкозы, аспарагина и КН2Р04). Колонки, закрытые чехлами из черной плотной бумаги, промывали дистиллированной водой с таким расчетом, чтобы периоды высокого и низкого увлажнения чередовались через 10 дней.
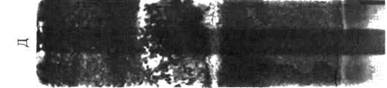
Через месяц на контакте горизонтов В и С появилось потемнение, которое стало постепенно распространяться вверх до середины горизонта В. Затем изменения прекратились. В конце второго месяца эксперимента в середине горизонта А2 появилась четко выраженная темная полоса (рис. 7а), которая постепенно стала увеличиваться по мощности и интенсивности окраски
|

|
|

|

|

|

|
(рис. 76). На исходе третьего месяца после очередного "сухого" периода, темная полоса стала исчезать сверху (рис. 7в) и, наконец, вовсе исчезла (рис. 7г). На ее месте остались бурые пятна. Последующее увлажнение почвенной колонки привело к образованию нового, но уже не сплошного и менее интенсивно окрашенного A2h в виде темных пятен и полос (рис. 7 д, е).
За год эксперимента произошло пятикратное образование и исчезновение гумусового горизонта в слое горизонта А2 насыпной колонки из материала реальных горизонтов подзолистой почвы. В последнем варианте на месте исчезнувших темных пятен образовались яркие ржаво-бурые пятна ожелезнения. Вероятно, при переувлажнении в анаэробных условиях происходит нисходящая миграция мелких частиц обугленной подстилки через рыхлую массу горизонта А2 до контакта с водоупором плотного горизонта В. После обсыхания в аэробных условиях происходит разложение органического вещества до конечных продуктов и на месте темного пятна остаются продукты его минерализации, которые окрашивают горизонт в бурые тона.
Эти предположения согласуются с литературными данными некоторых авторов. И.А. Соколов и Н.И. Белоусова (1964) отметили обилие древесных угольков в иллювиально-гумусо-вых горизонтах почв Камчатки. Де-Бано с соавторами (De Bano et al., 1970) объясняет формирование в некоторых почвах водоотталкивающего слоя с перемещением вниз по профилю гидрофобных органических веществ, образующихся в процессе горения.
Конечно, приведенные факты только косвенно подтверждают возможность существования в подзолистых почвах второго гумусового горизонта современного происхождения. Тем не менее, эта гипотеза имеет право на существование и объективное обсуждение наравне с господствующей реликтовой, тем более, что у последней нет прямых доказательств и их трудно воспроизвести в эксперименте.
Для обоснования гипотезы современной природы A2h необходимо аргументировать еще два важных положения:
- возможно ли в принципе образование и исчезновение второго гумусового горизонта в подзолистых почвах за сравнительно короткий период 100-150 лет, в течение которого происходит длительно производная сукцессия лесных экосистем;
- вполне ли адекватен описанный выше механизм формирования и исчезновения второго гумусового горизонта в почвах южной тайги природному процессу метаморфоза почвы.
Положительные ответы на эти вопросы могли бы существенно подкрепить гипотезу о современной природе A2h.
В литературе имеется много примеров фиксированного времени формирования почвенного профиля на породах известного возраста (крепостные стены, валы, курганы, молодые морены, отвалы и другое), которые показывают, что для превращения материнской породы в почвенный профиль достаточно одного столетия. Естественно предположить, что метаморфозы зрелых почв при значительном изменении внешних условий могут происходить и в более короткие периоды времени (Герасимов, 1968; Дюшофур,1970).
Судьба гумусовых веществ в почвенном профиле определяется соотношением двух противоположных процессов: гумификации разлагающейся некромассы и минерализации гумусовых веществ.
Время жизни различных фракций почвенного гумуса (характерное время) существенно различается. Фульвокислоты существуют 1-3 года, гуминовые кислоты 10—100 лет, гумины до 500 лет и больше. Даже гумины со временем полностью минерализуются и сменяются новыми. Во всех почвах происходит постоянный процесс естественного обновления почвенного гумуса. При этом каждая фракция имеет собственное характерное время обновления.
Приведенные примеры и собственный опыт полевых наблюдений позволяют предположить, что удаление лесного полога в зоне южной тайги как бы переносит подзолистую почву на период сукцессии в лесостепную зону. Изменившиеся гидротермические условия формируют новый биотоп, который стимулирует развитие высокозольных травянистых видов растений, активизирует почвенную микрофлору, увеличивает скорость биологического круговорота и способствует образованию в почвах гумат-ного гумуса.
Описанный выше механизм формирования второго гумусового горизонта можно считать частным случаем, поскольку он связан исключительно с катастрофическими сукцессиями темно-хвойных лесов южной тайги Причулымья, что не позволяет интерпретировать его как универсальный. Больше того, даже в этих конкретных условиях процесс формирования A2h складывается из двух составляющих, которые могут проявляться как совместно, так и в отдельности, образуя различные сочетания результатов.
Во-первых, после пожаров, в результате временного преобладания щелочной среды после озоления фитомассы, происходит щелочная экстракция естественного органического вещества
верхнего горизонта почвы и подстилки, перенос его в нижележащий А2 и осаждение на водоупоре горизонта В.
Во-вторых, во время лесных пожаров происходит обогащение толщи подзолистого горизонта древесным углем за счет цепной реакции обугливания сырой подстилки и корней при малом доступе кислорода. В почвенном разрезе обогащенный частичками древесного угля подзолистый горизонт выделяется темной окраской и фиксируется как A2h. Тем более, что стандартные методы анализа качества гумуса не позволяют отличить естественный гумус от смеси почвы с древесным углем.
В обоих случаях темноокрашенные органические вещества накапливаются на контакте горизонтов А2 и В, как за счет их разной водопроницаемости, так и вследствие высокой кислотности горизонта В, которая стимулирует осаждение гуминовых кислот из щелочных растворов.
Общеизвестную приуроченность почв со вторым гумусовым горизонтом к зоне южной тайги можно объяснить следующими соображениями. Во-первых, именно здесь особенно ярко выражен контраст микроклимата под пологом леса и на открытом пространстве. Следовательно удаление лесного полога здесь приводит к резкому изменению процесса почвообразования. На обширных таежных "еланях" среди подзолов часто встречаются дерново-подзолистые почвы с мощным темноокрашенным гумусовым горизонтом.
Во-вторых, зона южной тайги - это основной ареал вредоносности сибирского шелкопряда (Рожков, 1965), одного из существенных факторов уничтожения темнохвойных лесов на больших пространствах. По данным Н.Г. Коломийца (1957) крупные инвазии сибирского шелкопряда в районе Причулымья наблюдались в периоды: 1914-1917; 1920-1924; 1942-1946; 1952-1957 гг.
Многие исследователи растительности Причулымья отмечали существенную роль этого фактора катастрофических сукцессии в формировании растительного покрова южной тайги. По мнению некоторых исследователей (Дробов, 1909; Елизарьева, 1957; Куликов, 1966; Фуряев, 1970) лиственные и смешанные леса этого региона представляют различные стадии восстановления коренных темнохвойных насаждений после их гибели в результате инвазий сибирского шелкопряда и последующих пожаров.
Механизм цикличного метаморфоза заложен в перераспределении запаса минеральных элементов между ярусами экосистемы: сначала из почвы в фитомассу по мере созревания экосистемы и функционирования ее в фазе климакса, затем обратно после катастрофы. Виды-эдификаторы - главные потребители элементов минерального питания, выделяемых почвой в процессе
катаболизма. По мере увеличения фитомассы (анаболизм) в нее из почвы переходит все большее количество минеральных элементов, создавая в почве дефицит. По закону минимума из фитоценоза постепенно выпадают виды, чувствительные к дефициту минерального питания, а затем ослабевают и сами эдификаторы. Ослабление эдификатора провоцирует инвазии энтомовредите-лей и пожары, в результате которых происходит массовое отмирание фитомассы (некроболизм) и перемещение минеральных элементов в почву. Уничтожение лесного полога резко изменяет гидротермические условия на поверхности почвы, поэтому массированный приток некромассы компенсируется высокой активностью гетеротрофной биоты и способствует сохранению минеральных элементов в новом более устойчивом к минерализации гуматном гумусе.
Пятнистый почвенно-растительный покров южной тайги, включающий лесные поляны, участки жердняка, мелколесья, смешанных и чисто хвойных насаждений (до мертвопокровни-ков) представляют моментальный снимок различных фаз цикличного сукцессионного процесса. Вследствие того, что фазы имеют разное характерное время, а каждое пятно мозаики должно пройти полный цикл метаморфоза, создается реальная динамическая пестрота (мозаика) покрова. Следующий моментальный снимок через несколько лет отобразит уже иную картину мозаики с иным распределением в пространстве фаз сукцессии. Ускоренная съемка этого процесса могла бы создать реальное впечатление мерцания мозаики.
Мозаичность почвенно-растительного покрова характерна не только для таежных экосистем. Например, почвенно-растительный покров Прикаспийской низменности представляет собой очень пеструю мозаику, состоящую из ярко выраженных пятен, которые сочетаются в комплексы довольно однообразного состава. В пространстве меняются только количественные соотношения компонентов комплекса. На почвенных картах Прикаспийской низменности выделяются четырехчленные комплексы: каштановые, лугово-каштановые, солонцы и перерытые почвы сусликовых бугорков (бутонов). Лиманные солоди и солончаки образуют, как правило, крупные самостоятельные контуры.
Главной причиной своеобразной для региона пестроты почвенного покрова принято считать пестрый суффозионный рельеф территории, благодаря которому происходит перераспределение влаги редких дождей и маломощного снежного покрова на идеально ровной поверхности бессточной равнины. Именно дифференциация влаги в пространстве создает такую пестроту почвенного и растительного покрова.
Характерная для Прикаспийской низменности мозаика хорошо видна как на местности, так и на аэрофотоснимках. Эту картину принято считать неизменной и статичной, поскольку существует она много тысячелетий. Однако есть основания утверждать, что пятна мозаики довольно быстро меняются во времени и регулярно перемещаются в пространстве. Исследователь, зафиксировавший на карте или аэрофотоснимке мозаичный почвенный покров конкретного участка, через некоторое время, при повторной съемке может обнаружить смещение в пространстве комплексов и отдельных почв, иные очертания пятен и иные их сочетания в комплексе.
Инициаторы суффозионных процессов - грызуны-землерои и в первую очередь суслики. Высокая численность популяции суслика обыкновенного на территории Прикаспия и своеобразная морфология солонцового профиля - главный двигатель механизма естественной сукцессии почвенно-растительного покрова.
Суслик устраивает свое жилище в норе под сводами водонепроницаемого солонцового горизонта. В процессе рытья норы он выносит на поверхность рыхлый материал карбонатного и гипсового горизонтов, которые залегают непосредственно под иллювиальным горизонтом солонца. В сухом состоянии солонцовый горизонт представляет очень плотные, монолитные структурные отдельности (столбики, призмы, глыбы), разделенные широкими трещинами высыхания. При увлажнении весенними талыми водами или во время летних ливней, солонцовый горизонт сильно разбухает, становится вязким, гомогенным и не пропускает влагу в нижележащие горизонты почвы. Однако карбонатный и гипсовый горизонты находятся в сухом состоянии круглый год и жилище суслика гарантировано от затопления.
Разбухание солонцового горизонта происходит за счет пепти-зации коллоидов почвенного поглощающего комплекса, обогащенных катионами натрия. Основной прием мелиорации солонцов с целью улучшения их сельскохозяйственных свойств - это гипсование. Кальций гипса замещает натрий почвенного поглощающего комплекса, а вновь образованный легко растворимый сульфат натрия вымывается за пределы профиля.
В результате работы суслика по строительству нового жилища на поверхности солонца образуется конусовидный бугорок высотой 30-50 см и диаметром 1-2 м, состоящий из гипсоносной и карбонатной породы. Во время весеннего снеготаяния, когда Прикаспийская степь выглядит как рыбья чешуя из-за блюдец, падин и лиманов, заполненных водой, происходит активное выщелачивание сульфатов и карбонатов кальция из сусликового бугорка вниз до солонцового горизонта. Гипс активно взаимодей-
ствует с натрием почвенного поглощающего комплекса солонцового горизонта и разрушает его физико-химическую структуру.
Процесс естественного гипсования без участия человека вызывает "самомелиорацию" солонца, в которой суслик выполняет роль мелиоратора. Через несколько лет под влиянием процесса суффозии происходит просадка сусликового бугорка и его постепенное превращение в степное блюдце.
Вымывание токсичных солей (хлоридов и сульфатов) из материала бугорка талыми водами способствует его зарастанию сначала черной полынью, а затем злаками и разнотравьем. Под пологом растительности значительно активизируется процесс почвообразования. Высокие температуры и добавочное застойное увлажнение в короткий весенний период стимулируют развитие дернового процесса и обеспечивают формирование темноцветного профиля лугово-каштановых почв, характерных для блюдцеобразных понижений.
По мере увеличения глубины просадки и диаметра блюдец происходит их слияние. Соседние блюдца сливаются в падину, а падины, увеличиваясь в размерах и сливаясь с другими падинами, образуют обширные лиманы диаметром 1-2 км и больше. В лимане под влиянием застоя хорошо прогретых весенних талых вод, обедненных кислородом, активно развивается процесс осо-лодения почв, который гомогенизирует почвенный покров обширного замкнутого понижения.
В период временного весеннего переувлажнения верхних горизонтов почвы происходит даже кратковременное оглеение почвенного профиля с характерными морфологическими признаками лугово-болотных почв: творожистая структура, сизая окраска горизонтов, ржаво-бурые пятна. После испарения влаги, буквально в течение двух-трех недель, временный профиль с явными признаками лугово-болотной почвы превращается в профиль обычной лиманной солоди с характерными уже для нее морфологическими и физико-химическими признаками: серая окраска и слоеватая структура верхнего горизонта, оливковый цвет и плотная монолитная структура иллювиального горизонта.
Процесс осолодения почв со временем постепенно сменяется солонцовым и под его влиянием формируются каштановые солонцеватые почвы, солонцы и все начинается сначала. Под пологом солонцового горизонта поселяются суслики, их бугорки (бу-таны) мелиорируют солонец, который постепенно превращается в лугово-каштановую почву, а затем в лиманную солодь. Лиманная солодь превращается в солонец или каштановую солонцеватую почву. Все фазы этого процесса хорошо видны как на местности, так и на аэрофотоснимках. Только на местности лучше за-
метны начальные фазы: образование и просадка сусликовых бугорков, зарождение и слияние блюдец; а на аэрофотоснимке лучше видны последние фазы: слияние падин и зарождение солонцовых пятен на поверхности лимана.
Цикличность процесса проявляется примерно так же, как в случае сукцессий растительного покрова. Стадии строго следуют одна за другой до климакса, после чего начинается новый цикл.
В Читинской области нам удалось наблюдать как в течение трех лет после вырубки березового леса и распашки массива, светлоокрашенная (бурая) мерзлотная дерново-таежная почва превратилась в лугово-лесную почву с темно-серым профилем, похожим по окраске на профиль чернозема. Резкое различие гидротермических условий и баланса вещества под пологом леса и на открытом участке изменило ритмику функционирования экосистемы, которая преобразовала почвенный профиль и привела его в соответствие с новыми условиями экотопа.
Из сказанного выше можно сделать следующие выводы:
1. Все компоненты экосистемы функционируют синхронно с
динамикой и ритмикой гидротермических условий.
2. Многолетние флуктуации гидротермических условий в
рамках ограниченного диапазона создают устойчивую среду для
определенной ассоциации организмов, которые и формируют об
лик экосистемы.
3.Устойчивые отклонения гидротермических условий в пространстве или во времени образуют новый диапазон факторов среды, благоприятный для другой ассоциации организмов, которые формируют другой облик экосистемы. Если изменения наблюдаются в пространстве - это фиксируется топография экосистем, если во времени, то этот процесс протекает как метаморфоз.
4.Тенденция (тренды) изменений структуры экосистем в пространстве называется географией экосистем, во времени - эволюцией.
5.Антропогенные факторы могут значительно ускорить изменчивость экосистем во времени, поскольку оказывают прямое воздействие на их структуру. Изменчивость во времени неизбежно корректирует картину изменчивости в пространстве.
Приведенные выше примеры экогенетических сукцессии в таежных экосистемах Западной Сибири и полупустынных экосистемах Прикаспийской низменности демонстрируют механизм адаптации экосистем к сменившимся условиям среды. В таежных экосистемах инициатором сукцессии стал сибирский шелкопряд, а в Прикаспийской полупустыне суслик обыкновенный. В результате сукцессии изменились не только фитоценозы, но и педо-
ценозы, то есть вся экосистема прошла процедуру метаморфоза в течение сравнительно короткого времени в пределах 50 лет.
Нам удалось наблюдать более ускоренные метаморфозы почв в той же Прикаспийской низменности и в Восточном Забайкалье. Оказалось, что ранней весной, когда понижения рельефа Прикаспийской низменности заполнены талой водой в почвенном профиле создаются временные анаэробные условия и восстановительная обстановка. После испарения поверхностной влаги почвы понижений на протяжении 10—15 дней сохраняют в профиле восстановительные условия. Вскрытый разрезом профиль лиманной солоди был по всем морфологическим признакам похож на профиль лугово-болотной почвы. Через две недели эти признаки исчезли полностью и профиль лиманной солоди выглядит как обычно. Однако факт существования (пусть временного) лугово-болотной почвы на месте солоди зафиксирован и не подлежит сомнению. Почвоведы о нем не знают потому, что раньше мая в поле не выезжают, а факт был зафиксирован в середине апреля.
На Забайкальском мерзлотном стационаре мы изучали два основных типа почв: мерзлотно-таежные и мерзлотные лугово-лесные. Профиль первых светлый буроватый, а профиль вторых темно-серый почти черный. В первый год работы стационара был раскорчеван и распахан участок водораздельного березняка с мерзлотно-таежными почвами. Цвет пашни был светло бурый. Через три года цвет пашни стал темносерый, а профиль почвы приобрел признаки мерзлотной лугово-лесной почвы. За три года произошло изменение почвы на уровне типа. Факт ускоренного метаморфоза налицо. Таких примеров почвоведы-полевики знают множество, но несоответствие фактов господствующей теории до сих пор толкуется в пользу теории, вопреки фактам.
Заслуживает внимания еще один интересный и общеизвестный факт. Почвенная классификация разрабатывалась и в последствии много раз корректировалась на материалах почвенной съемки Европейской части России. В разгар картографических работ, связанных с составлением листов Почвенной карты СССР миллионного масштаба, сибирские почвоведы стали жаловаться московским руководителям работ на слишком частые несовпадения реальных признаков почв с диагностикой образцовых музейных профилей. Для решения накопившихся дискуссионных вопросов в Красноярске был созван 1-й съезд сибирских почвоведов, на котором академик И.П. Герасимов произнес крылатую фразу о самобытности сибирских почв. Эта фраза цитировалась в каждой статье, касающейся описания сибирских почв. Она давала право присваивать известные названия почвам с диагностическими признаками, отличными от европейских эталонов. Когда за-
вершился процесс полной инвентаризации почвенных ресурсов всей страны, то возник законный вопрос. Может быть уже настала пора говорить не о самобытности сибирских почв, а об уникальности почв Европейской России, площадь которой значительно меньше площади Сибири и Дальнего Востока. Но привычка к приоритету Москвы оставила этот вопрос открытым.
Главный критерий изменчивости и устойчивости почв и экосистем - это совокупность диагностических признаков, определяющая их принадлежность к определенному таксону классификации. Изменчивость в пространстве оценивается по трем категориям: пестрота, топография, география. Изменчивость во времени также делится на три категории: флуктуации, метаморфозы, эволюции.
Пестрота и флуктуации относятся к устойчивости экосистем в пространстве и во времени, они характеризуют изменчивость экосистемы в пределах диагностического диапазона. Экосистема меняется количественно, но остается в прежнем качестве, то есть в рамках прежнего таксона классификации. Только пестрота означает стабильность экосистемы в пространстве, а флуктуации -во времени.
Следующая по уровню категория изменчивости (топография и метаморфоз) означает переход экосистемы в другое качественное состояние, т.е. в другой таксон классификации. Топография характеризует изменчивость в пространстве, а метаморфоз - во времени. Другими словами, топография почв и экосистем указывает на каком расстоянии или в каких координатах пространства один таксон переходит в другой, а метаморфоз указывает в какой момент времени или через какое время данная почва или экосистема перейдет в другое качественное состояние. Топография почв и экосистем отражается на почвенных или экологических картах, а метаморфозы в экологических прогнозах и сценариях.
И наконец, высшие категории изменчивости (география и эволюция) представляют собой фундаментальные закономерности превращения таксонов в пространстве (география) и во времени (эволюция). Изменчивость в пространстве подчиняется закону природной зональности, а закон изменчивости во времени пока не сформулирован, его еще предстоит выявить и сформулировать. Закон природной зональности сформулирован, благодаря бурному развитию тематической картографии, которая документировала распределение в пространстве различных компонентов природных экосистем: почвы, растительность, геологические породы, климатические зоны и другое. Для формулирования закона изменчивости экосистем и почв во времени нужна такая же обширная и высокоточная база данных, характеризую-
щих динамику и ритмику процессов функционирования экосистем. Потребуется новая объективная система экологического мониторинга, включающая подсистемы измерения, оценки и прогноза состояния экосистем во времени, что-то вроде экологической кинематографии, способной фиксировать и документально отображать изменения во времени типичных динамических параметров экосистем и факторов внешней среды, контролирующих динамику и ритмику метаболизма экосистем.
Современные классификации природных экосистем оперируют устойчивыми во времени физиономичными (морфоструктур-ными) параметрами и поэтому позволяют объективно оценивать только изменчивость экосистем в пространстве методом картографирования. Для оценки изменчивости экосистем во времени статичные параметры не пригодны. Здесь нужна другая диагностика и классификация, основанная на динамических показателях, информативных во времени. Динамические параметры должны фиксировать количественные изменения метаболизма экосистем в суточном, сезонном, годовом и многолетнем циклах по принципу кинематографии.
На основе существующей классификации экосистем при помощи сравнительно-географического метода анализа реальных экологических ситуаций можно строить только предположения о возможной изменчивости экосистем во времени.
Ниже приведены два примера изменчивости почвенно-расти-тельного покрова в результате воздействия экстремальных естественных факторов зоогенного характера. В одном случае изменчивость почвенного покрова во времени стимулируется массовым размножением сибирского шелкопряда, а в другом - животными землероями - сусликами.
Восстановительные сукцессии фитоценозов известны давно и используются для оценки сравнительной устойчивости разных экосистем к различным факторам негативного воздействия. Инициаторами восстановительных сукцессий обычно были экстремальные экологические ситуации: пожары, инвазии энтомо-вредителей, распашка земель, обвалы, оползни, климатические аномалии и другое. Как правило, анализ сукцессии завершается после полного или частичного восстановления видового состава фитоценоза. Подразумевается, что дальнейшая судьба экосистемы прогнозируется однозначно - бесконечно долгое пребывание экосистемы в стадии климакса. Однако экосистема как и организмы, составляющие ее биоту, после достижения зрелости переходит во вторую фазу жизненного цикла неизбежного отмирания или некроболизма. В разных экосистемах некроболизм происходит по-разному, однако завершается он всегда одинаково - пол-
ной сменой фитоценоза с его длительным последующим восстановлением. Об изменениях почвы за этот период известно мало, поскольку почву принято считать инертной частью экосистемы, которая может изменяться только в масштабе геологического времени.
Такое положение противоречит определению экосистемы как целостной природной единицы, выполняющей функцию круговорота вещества и энергии или метаболизма. Если это так, то составляющие процесса метаболизма: анаболизм, осуществляемый фитоценозом, и катаболизм - педоценозом (почвой), должны функционировать синхронно, согласованно. А это значит, что почва обладает не только одним характерным временем геологического масштаба, но и другим, более коротким, адекватным характерному времени фитоценоза.
Практика земледелия и мелиорации дает множество примеров скоротечных изменений важных свойств почвы в течение 3-5 лет. И.А. Соколов и В.О. Таргульян (1976) 30 лет назад высказали идею о наличии у почвы двух характерных времен и предложили различать "почву-память" и "почву-момент". Это было первым признанием почвоведами наличия в почве параметров, меняющихся в годовом, сезонном и суточном циклах. Популярное среди почвоведов изучение на почвенных стационарах режимов тепла, влаги, дыхания, почвенных растворов осуществлялось без общей теоретической основы, как экзотика и сводились к статичной оценке типов температурного и водного режима почв как дополнительного диагностического признака.
Механизм адаптации.В схеме, представленной в таблице 3.2, отсутствуют значения выброса минеральных элементов, который обязательно происходит при переходе одной фракции в другую. В ней зафиксирована судьба оставшейся половины предыдущей фракции некромассы и ее дальнейшие превращения в почвенном профиле. Это объясняется тем, что именно из этих остатков формируется почвенный профиль и его горизонты. Эта видимая часть разлагающейся некромассы - лишь потенциальный запас вещества экосистемы. Вторая же часть, неучтенная нами половина некромассы, представляет актуальный пул минерального вещества, который почти полностью потребляется фитоценозом для выполнения функции анаболизма. Невостребованная фитоценозом малая часть этой минеральной массы утилизируется почвой, благодаря процессу гумификации. И только совсем небольшое количество минерального вещества выносится стоком из экосистемы.
Каждая фракция некромассы при переходе в следующую, выделяет свой набор и свое количество минеральных элементов.
В выделениях первых фракций разлагающейся некромассы, преобладают элементы в газообразной фазе, а в выделениях последних - минеральные соли (ионы) в форме водных растворов. Газы поступают в растения через листья, а соли (ионы) через корни. Наличие в почве одновременно всех фракций разлагающейся некромассы свидетельствует о том, что фитоценоз всегда имеет полный набор ЭМП, необходимых для фотосинтеза. Так должно быть при оптимальных сочетаниях условий среды. А в реальной природной обстановке все происходит гораздо сложнее.
Изменение гидротермических условий и аэрации почвенного профиля стимулирует изменение ритмики катаболизма экосистемы. При этом трансформация одних фракций ускоряется, других замедляется, третьих сохраняется в прежнем режиме, что нарушает геохимическую структуру выделений почвы. Такие изменения отражаются, прежде всего, на состоянии фитоценоза. Одни виды начинают испытывать дефицит элементов, другие избыток, третьи сохраняют статус кво. По этой причине начинается перестройка видовой структуры биологического сообщества. Так происходит адаптация биоценоза к изменениям условий среды. Этот механизм действует постоянно. Он контролирует видовую структуру сообщества таким образом, чтобы она с максимальной эффективностью обеспечивала выполнение функции анаболизма. Эффективность анаболизма выражается в количестве минеральных ресурсов, затраченных на производство единицы биомассы.
Классификация экосистем и их изменчивость во времени.Изменчивость и устойчивость экосистем определяется на основе сравнения диагностических признаков реальной экосистемы с идеальной, обозначенной в классификации или систематическом списке с заранее выбранными критериями разделения множества признаков на таксономические уровни.
Проблема объективной классификации природных экосистем стала актуальной после того, как начались работы по экологическому картографированию и стационарному изучению изменчивости экосистем во времени. Именно принадлежность к тому или иному таксону классификации дает основание для ответа на вопрос: изменилась экосистема качественно или осталась в прежнем состоянии с количественными поправками. Чем детальнее классификация, тем больше возможностей проследить степень изменчивости экосистем в пространстве и во времени.
Современные классификации экосистем базируются на визуальной органолептической оценке растительного покрова с указанием почвы и рельефа, что порождает массу накладок, пропусков и совпадений. Объективная классификация должна строить-
ся на основе фундаментальных представлений о сущности объекта исследований, в данном случае экосистемы. Поскольку за экосистемой устоялся приоритет функционирования, то и классификация экосистем должна строиться на основе функциональных параметров.
Для построения количественной классификации или систематики экосистем необходимо выбрать универсальный принцип, с помощью которого можно было бы составить всеобщую шкалу формальных признаков для диагностики экосистем. В основе универсального принципа должно быть представление о структуре экосистем как результате их функционирования в строго определенном диапазоне факторов внешней среды. Важнейшими естественными факторами являются: свет, тепло, влага. В связи с большой сложностью измерения и оценки световых характеристик (интенсивность и спектральный состав света, суточная, годовая, многолетняя продолжительность светового периода), в географии и экологии принято оперировать данными гидротермических условий (сочетание тепла и влаги). Кроме того, дефицит света испытывают только растения нижних ярусов лесных экосистем, но и в этом случае недостаток света может компенсироваться избытком С02. Для упрощения расчетов и моделирования функций, вполне допустимо отождествить гидротермические условия (сочетание тепла и влаги) с полной совокупностью естественных факторов среды, которые оказывают прямое воздействие на интенсивность или скорость процесса метаболизма и его составляющих: анаболизм, некроболизм, катаболизм. На экомассу и ее компоненты (биомасса, некромасса, минеральная масса) гидротермические условия воздействуют опосредованно через общую результативность процесса метаболизма и его составляющих.
Антропогенные факторы путем изъятия, привноса и трансформации массы вещества экосистемы оказывают прямое воздействие на ее структуру. В этом принципиальная разница факторов среды.
Детальное изучение структурно-функциональных связей между компонентами экосистемы и их реакций на изменения сочетаний факторов внешней среды, позволит адекватно моделировать процесс функционирования природных и аграрных экосистем и прогнозировать их возможные изменения при различных воздействиях. Кроме того, на основе таких знаний можно создать теорию управления механизмом функционирования природных и аграрных экосистем, выявить условия, при которых экосистема будет обладать максимальной продуктивностью и устойчивостью к негативным воздействиям.
Структура экосистемы определяется соотношением интен-сивностей процессов функционирования: анаболизма, некробо-лизма и катаболизма. Отсюда следует, что параметры структуры объективно отражают результативность функционирования экосистемы на определенный момент времени. Изменение внешних факторов меняет интенсивность функционирования, за которой следует изменение структуры. В результате такой цепочки изменений экосистема теряет одни признаки и приобретает другие, что позволяет перевести ее из одного таксона классификации в другой. При этом не важно в пространстве или во времени произошли эти изменения - однозначно, что экосистема изменилась и стала другой.
Наиболее обобщенная оценка состоянии экосистемы должна базироваться на ее общей массе - экомассе, которая представляет собой сумму биомассы, некромассы и минеральной массы (ми-нермассы). В свою очередь биомасса состоит из фитомассы, зоомассы и микробиомассы; некромасса состоит из опада, подстилки и гумуса; минермасса состоит из суммы газов, солей и коллоидов. Можно детализировать следующий уровень структуры, но и этого достаточно для построения принципов количественной классификации экосистем. Трудности возникают при первых же попытках измерения перечисленных параметров. Но это препятствие вполне преодолимо, преимущества количественной классификации способны компенсировать эти затраты.
Основываясь на количественных значениях экомассы и ее составляющих можно построить достаточно детальную классификацию экосистем. Для этого необходимо получить значения всех параметров для всег<
Дата добавления: 2020-10-01; просмотров: 691;
Поиск по сайту
Узнать еще
- E) Расчет структурных составляющих очага деформации с одним нейтральным сечением
- II. Химический состав природных воды
- II.2. Химический (молекулярный) состав нефтей и природных газов
- III этап. Составление программного кода
- А) Общие требования к составлению уравнений баланса электрической цепи
- А. Сеть и состав архивов
- А. Составление схемы технологического процесса
- Автоматизированные рабочие места: назначение, состав, виды обеспечения

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
