Побег и системы побегов
Побег, как и корень, - основной орган растения.Вегетативныепобеги в типичном случае выполняют функцию воздушного питания, но имеют ряд других функций и способны к разнообразным метаморфозам.Спороносные побеги (в том числе и цветок) специализированы как органырепродуктивные, обеспечивающие размножение.
Побег образуется верхушечной меристемой как единое целое и, поэтому, представляет собой единый орган того же ранга, что и корень. Однако по сравнению с корнем побег имеет более сложное строение. Вегетативный побег состоит из осевой части – стебля, имеющего цилиндрическую форму, илистьев – плоских боковых органов, сидящих на стебле. Кроме того, обязательной частью побега являютсяпочки– зачатки новых побегов, обеспечивающие нарастание побега и его ветвление, т.е. образование системы побегов. Главную функцию побега – фотосинтез – осуществляют листья; стебли – преимущественно несущие органы, выполняющие механическую и проводящую функции.
Главная черта, отличающая побег от корня, - его облиственность. Участок стебля, от которого отходит лист (листья) называется узел. Участки стебля между соседними узлами –междоузлия. Узлы и междоузлия повторяются вдоль оси побега. Таким образом, побег имеетметамерноестроение,метамером(повторяющимся элементом) побега являются узел с листом и пазушной почкой и нижележащее междоузлие (рис. 4.16).
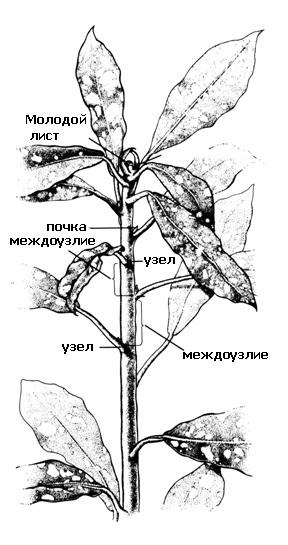
Рис. 4.16.Строение побега.
Первый побег растения – его главный побег, или побег первого порядка. Он образуется из зародышевого побега, заканчивающегосяпочечкой, которая формирует все последующие метамеры главного побега. По положению эта почка –верхушечная; пока она сохраняется, данный побег способен к дальнейшему росту в длину с образованием новых метамеров. Кроме верхушечной, на побеге образуютсябоковыепочки. У семенных растений они находятся в пазухах листьев и называютсяпазушными. Из боковых пазушных почек развиваютсябоковые побеги, и происходит ветвление, за счет которого увеличивается общая фотосинтезирующая поверхность растения. Формируетсясистема побегов, представленная главным побегом (побегом первого порядка) и боковыми (побегами второго порядка), а при повторении ветвления – боковыми побегами третьего, четвертого и последующих порядков. Побег любого порядка имеет свою верхушечную почку и способен к нарастанию в длину.
Почка– это зачаточный, еще не развернувшийся побег. Внутри почки заключена меристематическая верхушка побега – егоапекс (рис. 4.17).Апекс представляет собой активно работающий ростовой центр, который обеспечивает формирование всех органов и первичных тканей побега. Источником постоянного самообновления апекса являются инициальные клетки апикальной меристемы, сосредоточенные на кончике апекса. Вегетативный апекс побега, в отличие от всегда гладкого апекса корня, регулярно формирует на поверхности выступы, представляющие собой зачатки листьев. Гладким остается лишь самый кончик апекса, который называютконусом нарастанияпобега. Его форма сильно варьирует у разных растений и не всегда имеет вид конуса, вершинная часть апекса может быть низкой, полушаровидной, плоской или даже вогнутой.
Из вегетативных почек развиваются вегетативные побеги, состоящие из стебля, листьев и почек. Такая почка состоит из меристематической зачаточной оси, оканчивающейсяконусом нарастания, и зачаточных листьев разного возраста. Нижние листовые зачатки вследствие неравномерного роста загибаются внутрь и прикрывают верхние, более молодые, зачатки листьев и конус нарастания. Узлы в почке сближены, так как междоузлия еще не успели вытянуться. В пазухах листовых зачатков в почке уже могут быть заложены зачатки пазушных почек следующего порядка (рис. 4.17). Ввегетативно-генеративныхпочках заложен ряд вегетативных метамеров, а конус нарастания превращен в зачаточный цветок или соцветие.Генеративные, илицветочныепочки заключают в себе только зачаток соцветия или одиночного цветка, в последнем случае почку называютбутоном.
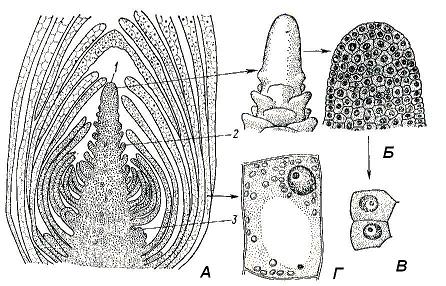
Рис. 4.17.Верхушечная почка побега элодеи:А – продольный срез; Б – конус нарастания (внешний вид и продольный срез); В – клетки апикальной меристемы; Г – паренхимная клетка сформировавшегося листа; 1 – конус нарастания; 2 – зачаток листа; 3 – зачаток пазушной почки.
Наружные листья почки часто видоизменяются в почечные чешуи, выполняющие защитную функцию и предохраняющие меристематические части почки от высыхания и резких перепадов температур. Такие почки называютсязакрытыми(зимующие почки деревьев и кустарников и некоторых многолетних трав).Открытыепочки не имеют почечных чешуй.
Кроме обычных, экзогенных по заложению, пазушных почек у растений часто образуются придаточные, илиадвентивныепочки. Они возникают не в меристематической верхушке побега, а на взрослой, уже дифференцированной части органа эндогенно, из внутренних тканей. Придаточные почки могут образоваться на стеблях (тогда они обычно расположены в междоузлиях), листьях и корнях. Придаточные почки имеют большое биологическое значение: они обеспечивают активное вегетативное возобновление и размножение тех многолетних растений, у которых они имеются. В частности, при помощи придаточных почек возобновляются и размножаютсякорнеотпрысковыерастения (малина, осина, осот, одуванчик).Корневые отпрыски– это побеги, развившиеся из придаточных почек на корнях. Придаточные почки на листьях образуются относительно редко. Если такие почки сразу дают маленькие побеги с придаточными корешками, которые отпадают от материнского листа и вырастают в новые особи, их называютвыводковыми(бриофиллум).
В сезонном климате умеренной зоны развертывание побегов из почек у большинства растений носит периодический характер. У деревьев и кустарников, а также у многих многолетних травянистых растений почки разворачиваются в побеги один раз в году – весной или в начале лета, после чего формируются новые зимующие почки с зачатками побегов будущего года. Побеги, вырастающие из почек за один вегетационный период, называют годичнымипобегами, илигодичными приростами. У деревьев они хорошо различаются благодаря образованиюпочечных колец– рубцов, которые остаются на стебле после опадения почечных чешуй. Летом у наших листопадных деревьев покрыты листьями годичные побеги только текущего года; на годичных побегах прошлых лет листьев уже нет. У вечнозеленых деревьев листья могут сохраняться на соответствующих годичных приростах 3-5 прошлых лет. В бессезонном климате за один год могут образоваться несколько побегов, разделенных небольшими периодами покоя. Такие побеги, образующиеся за один цикл роста, называютэлементарными побегами.
Почки, впадающие на некоторое время в состояние покоя, а затем дающие новые элементарные и годичные побеги, называют зимующимиилипокоящимися. По функции их можно назватьпочками регулярноговозобновления. Такие почки – обязательный признак любого многолетнего растения, древесного или травянистого, именно они обеспечивают многолетность существования особи. По происхождению почки возобновления могут быть и экзогенными (верхушечными или пазушными), и эндогенными (придаточными).
Если боковые почки не имеют периода ростового покоя и развертываются одновременно с ростом материнского побега, их называют почками обогащения. Развертывающиеся из нихпобеги обогащениясильно увеличивают (обогащают) общую фотосинтезирующую поверхность растения, а также общее число образуемых соцветий и, следовательно, семенную продуктивность. Побеги обогащения характерны для большинства однолетних трав и для ряда многолетних травянистых растений с удлиненными цветоносными побегами.
Особую категорию составляют спящие почки, очень характерные для лиственных деревьев, кустарников, кустарничков и ряда многолетних трав. По происхождению они, как и почки регулярного возобновления, могут быть пазушными и придаточными, но, в отличие от них, не разворачиваются в побеги в течение многих лет. Стимулом для пробуждения спящих почек служит обычно или повреждение основного ствола или ветви (пневая поросль после порубки у ряда деревьев), или естественное старение материнской системы побегов, связанное с затуханием жизнедеятельности нормальных почек возобновления (смена стволиков у кустарников). У некоторых растений из спящих почек на стволе образуются безлистные цветоносные побеги. Это явление носит названиекаулифлории и характерно для многих деревьев тропических лесов, например для шоколадного дерева. У гледичии из спящих почек на стволе вырастают пучки крупных разветвленных колючек – видоизмененных побегов (рис. 4.18).
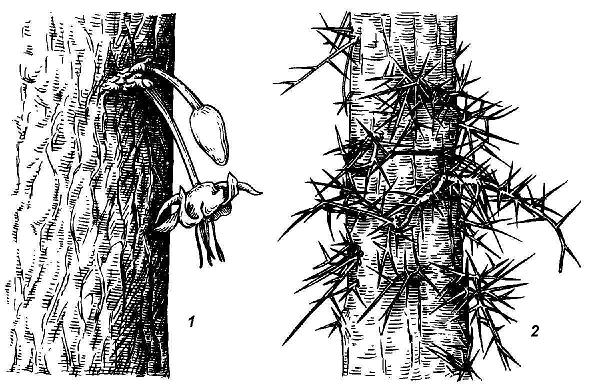
Рис. 4.18.Побеги из спящих почек:1 – каулифлория у шоколадного дерева; 2 – колючки у гледичии из разветвившихся спящих почек.
Направление роста побегов.Побеги, растущие вертикально, перпендикулярно поверхности земли, носят названиеортотропных. Горизонтально растущие побеги называютсяплагиотропными. Направление роста может меняться в процессе развития побега.
В зависимости от положения в пространстве различают морфологические типы побегов (рис. 4.19). Главный побег в большинстве случаев сохраняет ортотропный рост и остаетсяпрямостоячим. Боковые побеги могут расти в различном направлении, часто они образуют разной величины угол с материнским побегом. В процессе роста побег может изменить направление с плагиотропного на ортотропное, тогда он называетсяприподнимающимся, иливосходящим. Побеги с сохраняющимся в течение жизни плагиотропным ростом называютсястелющимися. Если они образуют в узлах придаточные корни, их называютползучими.
Ортотропный рост связан определенным образом со степенью развития механических тканей. При отсутствии хорошо развитых механических тканей в удлиненных побегах ортотропный рост невозможен. Но нередко растения, не обладающие достаточно развитым внутренним скелетом, все же растут вверх. Это достигается различными путями. Слабые побеги таких растений - лиан закручиваются вокруг какой-либо твердой опоры (вьющиесяпобеги), лазят с помощью различного рода шипиков, крючков, корней - прицепок (лазящиепобеги), цепляются с помощью усиков различного происхождения (цепляющиесяпобеги).
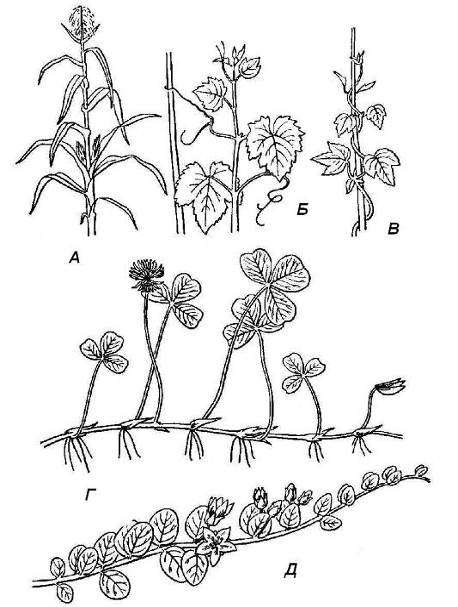
Рис. 4.19.Типы побегов по положению в пространстве: А – прямостоячий; Б – цепляющийся; В – вьющийся; Г – ползучий; Д – стелющийся.
Листорасположение.Листорасположение, илифиллотаксис– порядок размещения листьев на оси побега. Различают несколько основных типов листорасположения (рис. 4.20).
Спиральное, илиочередноелисторасположение наблюдается, когда на каждом узле расположен один лист, и основания последовательных листьев можно соединить условной спиральной линией.Двурядноелисторасположение можно рассматривать как частный случай спирального. При этом на каждом узле находится один лист, охватывающий широким основанием всю или почти всю окружность оси.Мутовчатоелисторасположение возникает, если на одном узле закладывается несколько листьев.Супротивноелисторасположение – частный случай мутовчатого, когда на одном узле образуются два листа, точно друг против друга; чаще всего такое листорасположение бываетнакрест супротивным, т.е. соседние пары листьев находятся во взаимно перпендикулярных плоскостях (рис. 4.20).
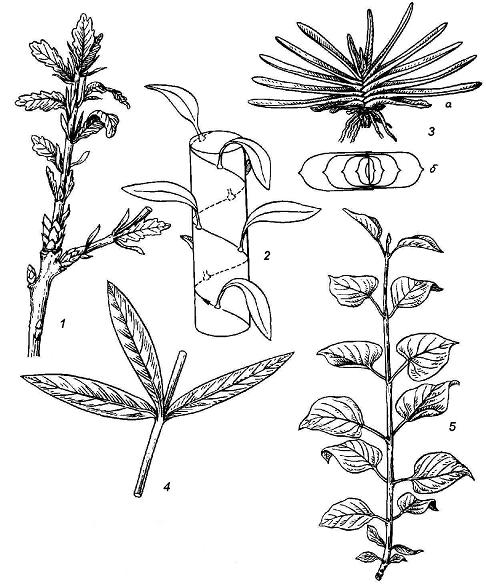
Рис. 4.20.Типы листорасположения: 1 – спиральное у дуба; 2 – схема спирального листорасположения; 3 – двурядное у гастерии (а – вид растения сбоку,б– вид сверху, схема); 4 – мутовчатое у олеандра; 5 – супротивное у сирени.
Порядок заложения листовых зачатков на апексе побега – наследственный признак каждого вида, иногда характерный для рода и даже целого семейства растений. Листорасположение взрослого побега определяется в первую очередь генетическими факторами. Однако в процессе развертывания побега из почки и его дальнейшего роста на расположение листьев могут оказывать влияние внешние факторы, главным образом, условия освещения и сила тяжести. Поэтому окончательная картина листорасположения может сильно отличаться от первоначальной и обычно приобретает ярко выраженный приспособительный характер. Листья располагаются так, что их пластинки оказываются в наиболее благоприятных в каждом конкретном случае условиях освещения. Наиболее ярко это проявляется в форме листовой мозаики, наблюдаемой на плагиотропных и розеточных побегах растений. При этом пластинки всех листьев располагаются горизонтально, листья не затеняют друг друга, а образуют единую плоскость, где нет просветов; более мелкие листья заполняют промежутки между крупными.
Типы ветвления побега.Ветвление – это образование системы осей. Оно обеспечивает увеличение общей площади соприкосновения тела растения с воздушной средой, водой или почвой. Ветвление возникло в процессе эволюции еще до появления органов. В простейшем случае верхушка главной оси вильчато ветвится и дает начало двум осям следующего порядка. Этоверхушечное, илидихотомическоеветвление. Верхушечным ветвлением обладают многие многоклеточные водоросли, а также некоторые примитивные растения, например плауны (рис. 4.21).
Для остальных групп растений характерен более специализированный боковойтип ветвления. При этом боковые ветви закладываются ниже верхушки главной оси, не затрагивая ее способности к дальнейшему нарастанию. При таком способе потенциальные возможности ветвления и образования систем органов гораздо более обширны и биологически выгодны.
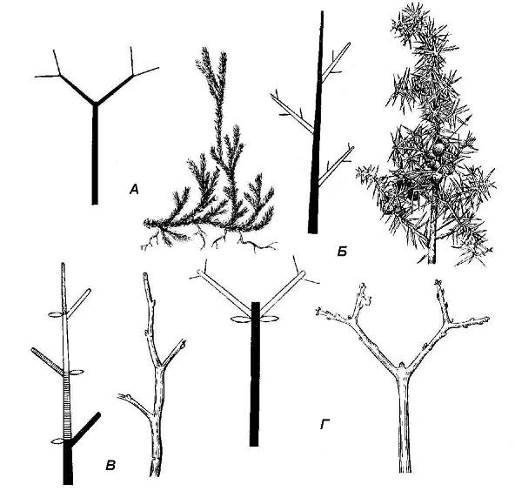
Рис. 4.21.Типы ветвления побега:А – дихотомическое (плаун); Б – моноподиальное (можжевельник); В – симподиальное по типу монохазия (черемуха); Г – симподиальное по типу дихазия (клен).
Различают два типа бокового ветвления: моноподиальноеисимподиальное (рис. 4.21). При моноподиальной системе ветвления каждая ось представляет собой моноподий, т.е. результат работы одной верхушечной меристемы. Моноподиальное ветвление характерно для большинства голосеменных и многих травянистых покрытосеменных растений. Большинство покрытосеменных растений, однако, ветвится по симподиальному типу. При симподиальном ветвлении верхушечная почка побега на определенном этапе отмирает или прекращает активный рост, но начинается усиленное развитие одной или нескольких боковых почек. Из них образуются побеги, замещающие побег, прекративший рост. Образовавшаяся при этом ось представляет собой симподий – составную ось, состоящую из осей нескольких последовательных порядков. Способность растений к симподиальному ветвлению имеет большое биологическое значение. В случае повреждения верхушечной почки рост оси будет продолжен боковыми побегами.
В зависимости от числа замещающих осей различают симподиальное ветвление по типу монохазия,дихазияиплейохазия. Ветвление по типу дихазия, илиложнодихотомическоеветвление характерно для побегов с супротивным листорасположением (сирень, калина).
У некоторых групп растений нарастание основных скелетных осей идет за счет одной или немногих верхушечных почек, боковые скелетные ветви совсем не образуются или образуются в очень малом числе. Древовидные растения такого типа встречаются главным образом в тропических областях (пальмы, драцены, юкки, агавы, саговники). Крона этих растений образована не ветвями, а крупными листьями, сближенными в розетку на верхушке ствола. Способность к быстрому разрастанию и захвату пространства, а также к восстановлению после повреждений у таких растений нередко отсутствует или слабо выражена. Среди деревьев умеренного климата такие неветвящиеся формы практически не встречаются.
Другая крайность – растения, слишком обильно ветвящиеся. Они представлены жизненной формой растений-подушек(рис. 4.22). Нарастание в длину побегов у этих растений крайне ограничено, но зато ежегодно образуется множество боковых веточек, расходящихся во всех направлениях. Поверхность системы побегов растения выглядит как бы подстриженной; некоторые подушки настолько плотны, что похожи на камни.
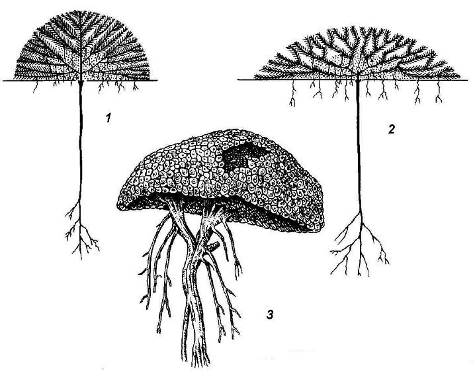
Рис. 4.22.Растения – подушки: 1, 2 – схемы строения растений-подушек; 3 – азорелла с острова Кергелен.
Очень сильно ветвятся представители жизненной формы перекати-поле, характерной для степных растений. Шаровидно разветвленная, очень рыхлая система побегов представляет собой огромное соцветие, которое после созревания плодов отламывается у основания стебля и перекатывается ветром по степи, рассеивая семена.
Специализация и метаморфозы побегов.У многих растений в пределах системы побегов наблюдается определенная специализация. Ортотропные и плагиотропные, удлиненные и укороченные побеги выполняют разные функции.
Удлиненными называют побеги с нормально развитыми междоузлиями. У древесных растений они называются ростовыми и располагаются по периферии кроны, определяя ее форму. Их главная функция – захват пространства, увеличение объема фотосинтезирующих органов.Укороченныепобеги имеют сближенные узлы и очень короткие междоузлия (рис. 4.23). Они формируются внутри кроны и поглощают проникающий туда рассеянный свет. Часто укороченные побеги у деревьев являются цветоносными и выполняют функцию размножения.
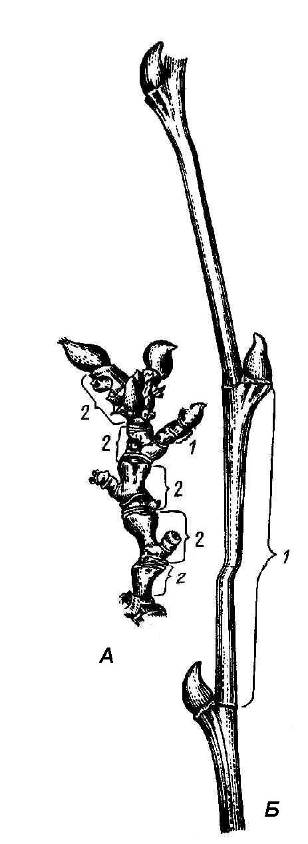
Рис. 4.23.Укороченный (А) и удлиненный (Б) побеги платана:1 – междоузлие; 2 – годичные приросты.
У травянистых растений обычно укороченные розеточныепобеги выполняют функцию многолетних скелетных и фотосинтезирующих, а удлиненные формируются в пазухах розеточных листьев и являются цветоносными (подорожник, манжетка, фиалки). Если пазушные цветоносы безлистны, их называютстрелками. Тот факт, что у древесных растений цветоносные побеги укороченные, а у травянистых – удлиненные, биологически хорошо объясним. Для успешного опыления соцветия трав должны быть приподняты над травостоем, а у деревьев даже укороченные побеги в кроне находятся в благоприятных для опыления условиях.
Примером специализации побегов могут служить многолетние осевые органы древесных растений – стволыисучьякроны. У листопадных деревьев годичные побеги теряют ассимиляционную функцию после первого вегетационного периода, у вечнозеленых – через несколько лет. Часть побегов после потери листьев отмирает целиком, но большинство остается в качестве скелетных осей, выполняющих опорную, проводящую и запасающую функции в течение десятков лет. Лишенные листьев скелетные оси известны под названиемсучьевистволов(у деревьев),стволиков (у кустарников).
В ходе приспособления к специфическим условиям среды обитания или в связи с резкой сменой функций побеги могут видоизменяться (метаморфизироваться). Особенно часто метаморфизируются побеги, развивающиеся под землей. Такие побеги утрачивают функцию фотосинтеза; они обычны у многолетних растений, где выступают в качестве органов переживания неблагоприятного периода года, запаса и возобновления.
Наиболее часто встречающимся подземным метаморфозом побега является корневище(рис. 4.24).Корневищем принято называть долговечный подземный побег, выполняющий функции отложения запасных питательных веществ, возобновления, а иногда и вегетативного размножения. Корневище формируется у многолетних растений, как правило, не имеющих во взрослом состоянии главного корня. По положению в пространстве оно может бытьгоризонтальным,косымиливертикальным. Корневище обычно не несет зеленых листьев, но, будучи побегом, сохраняет метамерную структуру. Узлы выделяются либо по листовым рубцам и остаткам сухих листьев, либо по живым чешуевидным листьям, также в узлах располагаются пазушные почки. По этим признакам корневище легко отличить от корня. Как правило, на корневище формируются придаточные корни; из почек вырастают боковые ответвления корневища и надземные побеги.
Формируется корневище или изначально как подземный орган (купена, вороний глаз, ландыш, черника), или сначала как надземный ассимилирующий побег, который затем погружается в почву с помощью втягивающих корней (земляника, медуница, манжетка). Корневища могут нарастать и ветвиться моноподиально (манжетка, вороний глаз) или симподиально (купена, медуница). В зависимости от длины междоузлий и интенсивности нарастания, различают длинныеикороткиекорневища и, соответственно,длиннокорневищныеикороткокорневищныерастения.
При ветвлении корневищ образуется куртинанадземных побегов, связанных участками системы корневищ. Если связующие части разрушаются, побеги обособляются, и происходит вегетативное размножение. Совокупность новых особей, образовавшихся вегетативным путем, называютклоном. Корневища характерны преимущественно для травянистых многолетников, но встречаются и у кустарников (бересклет) и кустарничков (брусника, черника).
Близки к корневищам подземныестолоны– недолговечные тонкие подземные побеги, несущие недоразвитые чешуевидные листья. Столоны служат для вегетативного размножения, расселения и захвата территории. Запасные питательные вещества в них не откладываются.
У некоторых растений (картофель, земляная груша) к концу лета из верхушечных почек столонов образуются клубни(рис. 4.24). Клубень имеет шаровидную или овальную форму, стебель сильно утолщен, в нем откладываются запасные питательные вещества, листья редуцируются, в их пазухах формируются почки. Столоны отмирают и разрушаются, клубни перезимовывают, и на следующий год дают начало новым надземным побегам.
Клубни не всегда развиваются на столонах. У некоторых многолетних растений клубневидно разрастается и утолщается основание главного побега (цикламен, капуста кольраби) (рис. 4.24). Функции клубня – запас питательных веществ, переживание неблагоприятного периода года, вегетативное возобновление и размножение.
У многолетних трав и полукустарничков с хорошо развитым стержневым корнем, сохраняющимся в течение всей жизни, образуется своеобразный орган побегового происхождения, называемый каудексом. Вместе с корнем он служит местом отложения запасных веществ и несет множество почек возобновления, часть из которых могут быть спящими. Каудекс обычно бывает подземным и образуется из коротких оснований побегов, погружающихся в почву. От коротких корневищ каудекс отличается способом отмирания. Корневища, нарастая верхушкой, постепенно отмирают и разрушаются на более старом конце; главный корень не сохраняется. Каудекс разрастается в ширину, с нижнего конца он постепенно переходит в долгоживущий утолщающийся корень. Отмирание и разрушение каудекса и корня идет от центра к периферии. В центре образуется полость, а затем он может разделиться продольно на отдельные участки –партикулы. Процесс разделения особи стержнекорневого растения с каудексом на части называютпартикуляцией. Каудексовых растений много среди бобовых (люпин, люцерны), зонтичных (бедренец, ферула), сложноцветных (одуванчик, полыни).
Луковица– это, как правило, подземный побег с очень коротким уплощенным стеблем –донцеми чешуевидными мясистыми сочными листьями, запасающими воду и растворимые питательные вещества, главным образом сахара. Из верхушечной и пазушных почек луковиц вырастают надземные побеги, на донце образуются придаточные корни (рис. 4.24). Таким образом, луковица – типичный орган вегетативного возобновления и размножения. Луковицы наиболее характерны для растений из семейств лилейных (лилии, тюльпаны), луковых (луки) и амариллисовых (нарциссы, гиацинты).
По структуре луковицы очень разнообразны. В одних случаях запасающие чешуи луковицы представляют собой только видоизмененные листья, не имеющие зеленых пластинок (лилия саранка); в других – это подземные влагалища зеленых ассимилирующих листьев, которые утолщаются и остаются в составе луковицы после отмирания пластинок (лук репчатый). Нарастание оси луковицы может быть моноподиальным (подснежник) или симподиальным (гиацинт). Наружные чешуи луковицы расходуют запас питательных веществ, подсыхают и играют защитную роль. Число луковичных чешуй варьирует от одной (чеснок) до нескольких сотен (лилии).
В качестве органа возобновления и запаса луковица приспособлена главным образом к климатам средиземноморского типа – с достаточно мягкой влажной зимой и с очень жарким засушливым летом. Она служит не столько для благополучной перезимовки, сколько для переживания жесткой летней засухи. Запасание воды в тканях луковичных чешуй происходит за счет образования слизей, способных удерживать большое количество воды.
Клубнелуковицавнешне напоминает луковицу, но ее чешуевидные листья не являются запасающими; они сухие и пленчатые, а запасные вещества откладываются в утолщенной стеблевой части (шафран, гладиолус).

Рис. 4.24.Подземные метаморфозы побега: 1, 2, 3, 4 – последовательность развития и строение клубня картофеля; 5 – клубень цикламена; 6 – клубень кольраби; 7 – луковички лилии тигровой; 8 – луковица лука репчатого; 9 – луковица лилии; 10 – участок длинного корневища пырея ползучего.
Видоизменяться могут не только подземные, но и надземные побеги растений (рис. 4.25). Довольно часто встречаютсянадземныестолоны. Это плагиотропные недолговечные побеги, функция которых вегетативное размножение, расселение и захват территории. Если столоны несут зеленые листья и участвуют в процессе фотосинтеза, их называютплетями(костяника, живучка ползучая). У земляники столоны лишены развитых зеленых листьев, стебли у них тонкие и хрупкие, с очень длинными междоузлиями. Такие более высокоспециализированные для функции вегетативного размножения столоны называютусами.
Сочными, мясистыми, приспособленными для накопления воды могут быть не только луковицы, но и надземные побеги, обычно у растений, живущих в условиях недостатка влаги. Водозапасающими органами могут быть листья или стебли, иногда даже почки. Такие сочные растения носят название суккулентов.Листовые суккуленты запасают воду в тканях листьев (алоэ, агавы, толстянки, родиола, или золотой корень). Стеблевые суккуленты характерны для американского семейства кактусовых и африканских молочайных. Сочный стебель выполняет водозапасающую и ассимилирующую функцию; листья редуцированы или превращены в колючки (рис. 4.25, 1).У большинства кактусов стебли колонновидные или шаровидные, листьев на них совсем не образуется, но узлы хорошо заметны по расположению пазушных побегов –ареол, имеющих вид бородавок или удлиненных выростов с колючками или пучками волосков. Превращение листьев в колючки уменьшает испаряющую поверхность растения и защищает от поедания животными. Примером метаморфоза почки в суккулентный орган –кочанслужит культурная капуста.
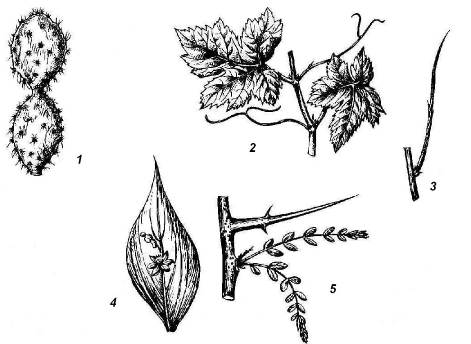
Рис. 4.25.Надземные метаморфозы побега:1 – стеблевой суккулент (кактус); 2 – усики винограда; 3 – безлистный фотосинтезирующий побег дрока; 4 – филлокладий иглицы; 5 – колючка гледичии.
Колючкикактусов имеют листовое происхождение. Листовые колючки нередко встречаются и у несуккулентных растений (барбарис) (рис. 4.26, 1).У многих растений колючки имеют не листовое, а стеблевое происхождение. У дикой яблони, дикой груши, жостера слабительного в колючки метаморфизируются укороченные побеги, имеющие ограниченный рост и оканчивающиеся острием. Вид жесткой одревесневшей колючки они приобретают после опадения листьев. У боярышника (рис.4.26, 3)колючки, образующиеся в пазухах листьев, совершенно безлистны с самого начала. У гледичии (рис. 4.25, 5) мощные разветвленные колючки образуются на стволах из спящих почек. Образование колючек любого происхождения, как правило, есть результат недостатка влаги. При выращивании многих колючих растений в искусственной влажной атмосфере они утрачивают колючки: вместо них вырастают нормальные листья (верблюжья колючка) или облиственные побеги (английский дрок).
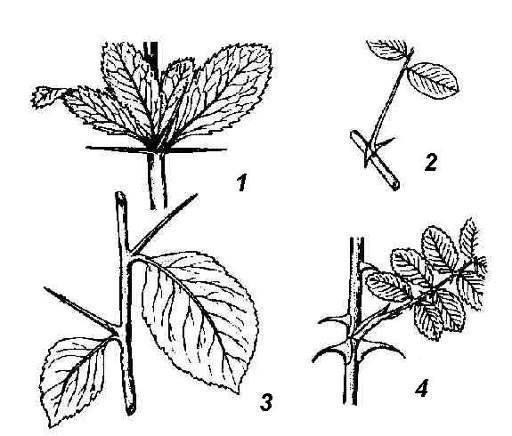
Рис. 4.26. Колючки различного происхождения: 1 – листовые колючки барбариса; 2 – колючки белой акации, видоизменение прилистников; 3 – колючки боярышника побегового происхождения; 4 – шипы – эмергенцы шиповника.
Побеги ряда растений несут шипы. Шипы отличаются от колючек меньшими размерами, это выросты – эмергенцы – покровной ткани и тканей коры стебля (шиповник, крыжовник) (рис. 4.26, 4).
Приспособление к недостатку влаги очень часто выражается в ранней утрате, метаморфозе или редукции листьев, теряющих основную функцию фотосинтеза. Это компенсируется тем, что роль ассимилирующего органа берет на себя стебель. Иногда такой ассимилирующий стебель безлистного побега остается внешне неизмененным (испанский дрок, верблюжья колючка) (рис. 4.25, 3).Дальнейшим шагом в указанной смене функций является формирование таких органов, какфиллокладииикладодии. Это уплощенные листоподобные стебли или целые побеги. На побегах иглицы (рис. 4.25, 4), в пазухах чешуевидных листьев, развиваются плоские листовидные филлокладии, имеющие, подобно листу, ограниченный рост. На филлокладиях образуются чешуевидные листья и соцветия, чего никогда не бывает на нормальных листьях, значит, филлокладий соответствует целому пазушному побегу. Мелкие, игольчатые филлокладии образуются у спаржи в пазухах чешуевидных листьев основного скелетного побега. Кладодии – это уплощенные стебли, сохраняющие, в отличие от филлокладиев, способность к длительному росту.
Для некоторых растений характерно видоизменение листьев или их частей, а иногда целых побегов в усики, которые закручиваются вокруг опоры, помогая тонкому и слабому стеблю сохранять вертикальное положение. У многих бобовых в усики превращается верхняя часть перистосложного листа (горох, горошки, чина). В других случаях в усики превращаются прилистники (сассапариль). Очень характерные усики листового происхождения формируются у тыквенных, причем можно видеть все переходы от нормальных листьев к полностью метаморфизированным. Усики побегового происхождения можно наблюдать у винограда (рис. 4.25, 2),пассифлоры и ряда других растений.
Стебель
Стебель представляет собой ось побега, состоящую из узлов и междоузлий. Основные функции стебля – опорная (несущая) и проводящая. Стебель осуществляет связь между корнями и листьями. В многолетних стеблях обычно откладываются запасные питательные вещества. Молодые стебли, имеющие под эпидермой хлоренхиму, активно участвуют в фотосинтезе.
Стебель обычно имеет цилиндрическую форму и характеризуется радиальной симметрией в расположении тканей. Однако в поперечном сечении он может быть не только округлым, но и угловатым –трех-,четырех-илимногогранным,ребристым,бороздчатым, иногда совершенно плоским,сплюснутым, или несущим выступающие плоские ребра -крылатым(рис. 4.27).
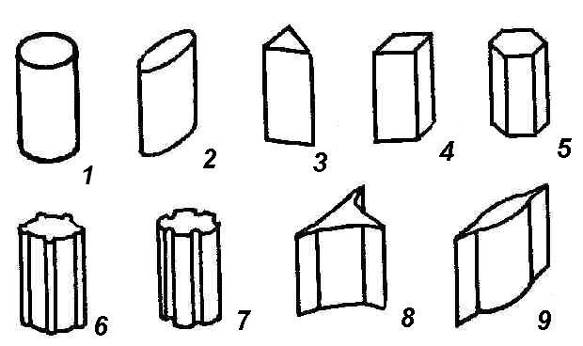
Рис. 4.27. Типы стеблей по форме поперечного сечения: 1 – округлый; 2 – сплюснутый; 3 – трехгранный; 4 – четырехгранный; 5 – многогранный; 6 – ребристый; 7 – бороздчатый; 8, 9 – крылатые.
Стебли древесных и травянистых растений резко различаются по продолжительности жизни. Надземные побеги трав сезонного климата живут, как правило, один год; продолжительность жизни побегов определяется продолжительностью жизни стебля. У древесных растений стебель существует много лет.
Анатомическое строение стеблясоответствует его главным функциям. В стебле развита сложная система проводящих тканей, которая связывает в единое целое все органы растения; наличие механических тканей обеспечивает выполнение опорной функции. Стебель, как и в целом побег, представляет собой «открытую» систему роста, он длительное время нарастает и на нем возникают новые органы.
Ткани стебля образуются в результате деятельности сложной системы меристем: верхушечных, боковых и вставочных (рис. 4.28).Первичная структура складывается в результате работы первичных меристем. Инициальные клеткиапикальноймеристемы сосредоточены в конусе нарастания побега. На апексе побега с правильной периодичностью возникают листовые зачатки, что приводит к раннему вычленению узлов, а развитие междоузлий запаздывает. Часто рост междоузлий и развитие в них постоянных тканей продолжаются длительное время за счет работы остаточныхинтеркалярныхмеристем, которые сохраняются у оснований молодых междоузлий. Хорошим примером такого интеркалярного (вставочного) роста может служить стебель злаков, у которого апикальная меристема очень рано расходуется на образование соцветия, и быстрое вытягивание побега обязано именно вставочному росту.
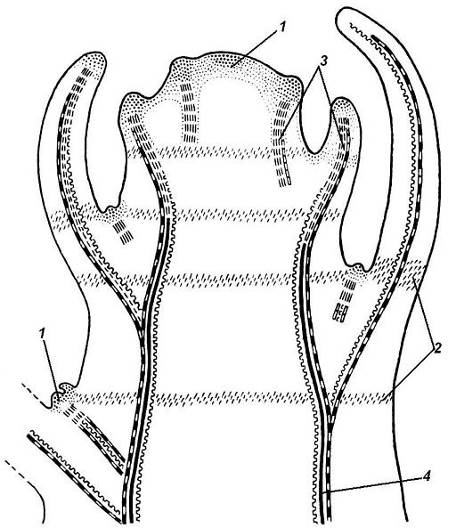
Рис. 4.28.Схема распределения меристем в стебле: 1 – апикальная меристема; 2 – интеркалярная меристема; 3 – прокамбий; 4 – камбий.
Самый наружный слой клеток апекса становится протодермой, из которой развивается эпидерма – покровная ткань будущего листа и стебля. На уровне первых листовых бугорков в апикальной меристеме обозначаются тяжи более узких и длинных клеток – этопрокамбий, дающий начало первичным проводящим тканям. Прокамбий может возникнуть в виде отдельных пучков или сплошного кольца. По мере дальнейшего роста прокамбий распространяется как в растущий листовой зачаток, так и в стебель, образуя основу будущей проводящей системы побега, связывающей листья и стебли. Остальная часть апекса занятаосновной меристемой, из которой в дальнейшем формируются паренхимные запасающие и ассимилирующие ткани, а также первичные механические ткани. Основная меристема, находящаяся между протодермой и прокамбием, превращается в первичную кору стебля, а из основной меристемы, располагающейся в центре, образуется сердцевина.
Первичная структура стебля у споровых и однодольных растений сохраняется в течение всей жизни. У голосеменных и двудольных внутри прокамбия возникает камбий, который откладывает вторичные проводящие ткани, что приводит к вторичному утолщению стебля.
Первичное строение стебля.В стебле, имеющем первичное строение, как и в корне, различаютпокровную ткань,первичную коруистелу (осевой, илицентральный цилиндр) (рис. 4.29).
Покровной тканьюявляетсяэпидерматипичного строения. В составпервичной корывходит основная паренхима, а также механические, выделительные и некоторые другие ткани. Из механических тканей чаще встречаетсяколленхима, она образует либо сплошной цилиндр, либо имеет вид отдельных тяжей, располагающихся обычно вдоль выступов – ребер стебля (рис. 4.29). Сразу под колленхимой или эпидермой, если колленхима отсутствует, в условиях, благоприятных для фотосинтеза, располагаетсяхлоренхима. Она может образовывать с колленхимой или склеренхимой чередующиеся полосы, проходящие вдоль стебля. Граница между корой и стелой выражена гораздо менее че
Дата добавления: 2016-05-30; просмотров: 14683;
Поиск по сайту
Узнать еще
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- I.2. Антигены системы АВ0. Генетика. Структура
- I.2.1 ПЕРВЫЙ ЗАКОН НЬЮТОНА. ИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА.
- I.2.1 ПОЛНАЯ И ВНУТРЕННЯЯ ЭНЕРГИЯ СИСТЕМЫ. ТЕПЛОТА И РАБОТА
- I.3. Антитела системы АВ0
- I.5.4 НЕИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА. СИЛЫ ИНЕРЦИИ
- II. ЭЛЕКТРИЧЕСКИЙ ДИПОЛЬ. ДИПОЛЬНЫЙ МОМЕНТ СИСТЕМЫ ЭЛЕКТРИЧЕСКИХ ЗАРЯДОВ
- III. Избирательные системы.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
