Использование неорганических доноров электронов
В клетках большинства организмов при дыхании донорами электронов являются органические вещества, которые одновременно выполняют роль источников углерода. Однако среди прокариот существуют группы почвенных и водных бактерий, способные использовать в качестве доноров электронов неорганические восстановленные вещества: элементарную серу, молекулярный водород, окись углерода, а также ионы аммония, нитрита, сульфита, тиосульфата, сульфида, двухвалентного железа. Эти бактерии впервые обнаружил и описал С.Н. Виноградский, назвав способ их метаболизма «хемосинтезом». Данные бактерии по типу питания относят к группе хемолитоавтотрофов: в качестве источника энергии они используют энергию химических связей химических соединений, донором электронов служат неорганические вещества, а источником углерода — СО2. 2
У литотрофных бактерий количество цитохромов в дыхательной цепи обычно в несколько раз превышает таковое у органотрофов. Связано это с особенностями используемых доноров электронов, которые в большинстве случаев характеризуются более положительными значениями Е0’, чем у NAD, т. е. не могут передавать свои электроны на NAD+ . Включение электронов, отщепляемых от большинства неорганических субстратов, в дыхательную цепь осуществляется чаще всего на уровне флавина или цитохромов. Например, первичным акцептором электронов при окислении ионов Fe2+ является цитохром с, а при окислении ионов NO2- — цитохром а. Поэтому у железобактерий и бактерий-нитрификаторов в электронтранспортной цепи функционируют лишь III-й и IV-й либо даже только IV-й комплексы. Соответственно, число событий фосфорилирования ADP уменьшается до двух или одного против трех, имеющих место у органотрофных бактерий при переносе электронов на NAD+ . Исключение представляют водородные бактерии: для Н2 значение Е’0=-0,42 В, и при окислении молекулярного водорода под действием О2 происходит перенос электронов через все три участка сопряжения, что приводит к синтезу 3 молекул АТР.
Существование укороченной дыхательной цепи у большинства хемолитоавтотрофов является причиной неэффективного запасания энергии в ходе дыхания, что вызывает необходимость перерабатывать огромные количества субстрата. В результате эти бактерии очень медленно растут, например, промежутки между двумя делениями клетки у бактерий-нитрификаторов достигают 5— 10 ч. Кроме этого, у данных бактерий возникает дефицит NADH в клетках, поскольку большинство неорганических доноров электронов не в состоянии восстанавливать NAD+ . Чтобы справиться с этой проблемой, у хемолитотрофов выработался механизм обратного транспорта электронов по компонентам дыхательной цепи. Такой обращенный поток электронов приводится в движение гидролизом АТР.
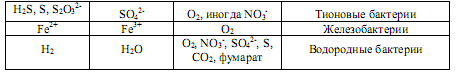
В табл. 12.3 охарактеризованы некоторые процессы использования неорганических доноров электронов. Следует отметить, что среди хемолитоавтотрофов встречаются как облигатные, так и факультативные литотрофы. Последние способны также к хемоорганогетеротрофному способу питания.
Окисление неорганических соединений может происходить как в аэробных (с участием О2), так и в анаэробных условиях, т. е. в процессе анаэробного дыхания (табл. 12.3).
Хемолитоавтотрофы способны фиксировать СО2, и большинство из них использует для этой цели цикл Кальвина.

Дата добавления: 2016-05-30; просмотров: 2191;
Поиск по сайту
Узнать еще
- II. Судовождение с использованием лоцманского метода и графического счисления пути судна.
- А) Строительство гнезд (или использование других убежищ для икры).
- Автоматизированная информационная система организации перевозок грузов по безбумажной технологии с использованием электронной накладной (АИС ЭДВ)
- Автоматизированный асинхронный электропривод с использованием синхронных электромашинных преобразователей частоты.
- Автоматизированный электропривод с использованием ПЧ с ШИР.
- Автоматическая локомотивная сигнализации и ее виды. Общий принцип работы АЛС с использованием рельсовых цепей
- Автомодельного ламинарного течения жидкости в трубе с использованием системы дифференциальных уравнений, описывающих течение жидкости в трубе
- Алгоритм расчета структуры изображения с использованием функции размытия линии.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
