Использование гидрохимических характеристик для выявления динамики вод океана
Требования к гидрохимическим работам в последнее время значительно возросли. На современном уровне требуется не только дать ту или иную пространственную картину распределения данного элемента, но и выявить основные процессы, контролирующие поведение этого элемента в океане.
Более того, следует указать относительную значимость каждого процесса, его -скорость и направление в данном районе океана. К сожалению, материал полевых наблюдений, которым сейчас располагают гидрохимики, почти не оставляет возможностей для такого анализа.
Основная масса гидрохимических исследований в океане проводилась только с географическими целями в период, когда большая часть Мирового океана представляла «белое пятно». Теперь потребовалось выяснить не только основные закономерности пространственного распределения гидрохимических параметров, но и понять процессы, определяющие их поведение.
Понятно, что перед гидрохимиками возникла дилемма: или начать заново исследование океана, уделяя основное внимание изучению процессов био- и физико-химических превращений элементов, или найти возможность использовать огромный фактический материал, накопленный почти за вековую историю океанологии, для решения новых задач. Решение в данном случае, как и следовало ожидать, будет компромиссным, а не альтернативным, т. е. потребуются новые исследования для познания химических процессов в океане, которые в свою очередь помогут по-новому прочитать и осмыслить имеющийся материал стандартных гидрохимических наблюдений.
Огромную работу в этом направлении проделали Харвей (1948), Редфилд (1934), X. Р. Сейвелл (1937), С. В. Бруевич (1937) и Б. А. Скопинцев (1950). Своими работами по исследованию процессов ассимиляции и регенерации биогенных веществ они заложили фундамент морской биогидродимии. Наиболее значительным достижением этих работ следует признать определение количественных взаимосвязей между потреблением кислорода и регенерацией фосфатов, кремния и соединений азота.
Первый шаг по использованию результатов этих биохимических работ для анализа гидрохимических данных сделал А. Редфилд (1942). Он ввел расчетный параметр преформ- фосфаты (preformed phosphate) и разделил величину наблюдаемой концентрации фосфатов на: а) первоначально образованные фосфаты, т. е. концентрация фосфатов, которая сформировалась в районе образования данной водной массы, б) фосфаты, высвобождаемые при разложении и окислении органического вещества, сформированного в том же районе, и в) фосфаты, регенерируемые органическим веществом, опускающимся из поверхностного слоя по мере движения вод.
Таким образом, он разделил величину концентрации фосфатов, наблюдаемую in situ, на генетические компоненты. Трудно переоценить значение этой работы. Для сравнения можно сказать, что для такой характеристики, как соленость, наиболее детально изученной в океанологии, вряд ли даже в будущем представится возможность выделить ее генетические компоненты.
Вслед за этой работой появились исследования в том же направлении. Т. Томпсон (1958) ввел понятие преформ-нитратов и преформ-силикатов, а Д. Дж. Рочфорд (1961) впервые построил карту пространственного распределения преформ- фосфатов для юго-восточной части Индийского океана. Из отечественных исследователей можно сослаться только на работу Д. А. Сметанина (1957).
Автором данной статьи уже использован расчет преформ- фосфатов при обработке материала для монографии «Химия Тихого океана» (В. В. Сапожников и В. В. Мокиевская, 1965). Разрешающие возможности данного метода и его практическое применение разобраны на примере сравнительного изучения Тихого и Атлантического океанов (В. В. Сапожников, 1965). В этой же работе указаны условия применимости данного метода для анализа гидрохимических данных.
В данной работе проведен расчет преформ-фосфатов на девяти разрезах в Индийском океане. Разрезы выбраны таким образом, чтобы приблизительно равномерно покрыть всю акваторию океана. По результатам расчетов построены карты распределения преформ-фосфатов и дефицита кислорода на 1000 м. Горизонт 1000 м выбран потому, что на большей части океана именно на этой глубине распространяется промежуточная субантарктическая водная масса, которая характеризуется максимальными концентрациями преформ-фосфатов. В юго-восточной части океана изолинии проведены по данным Д. Дж. Рочфорда (1961).
Меридиональные разрезы, на которых дано распределение температуры, солености, фосфатов, кислорода, преформ-фосфатов и дефицита кислорода (рис. 22—27), обнаруживают поразительное сходство в основных деталях, отражающих макроструктуру океана.
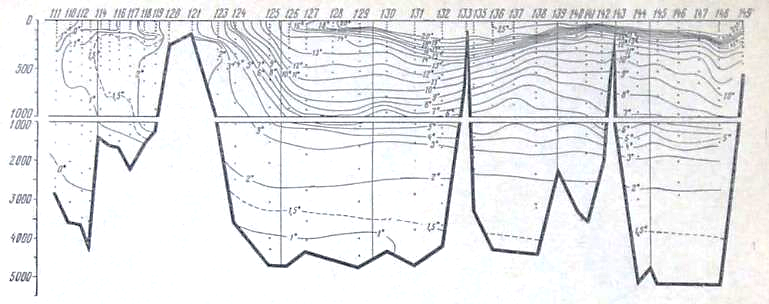
Рис. 22. Распределение температуры на разрезе море Девиса — мыс Гвардафуй
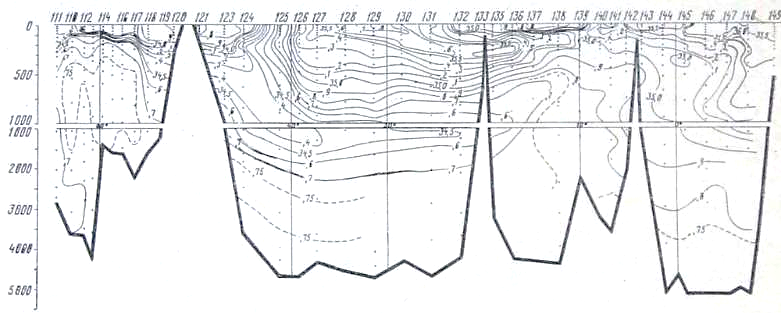
Рис. 23. Распределение солености на разрезе море Девиса — мыс Гвардафуй
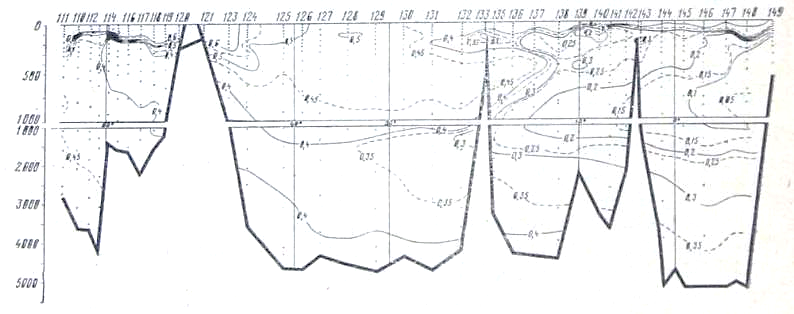
Рис. 24. Распределение кислорода на разрезе море Девиса — мыс Гвардафуй
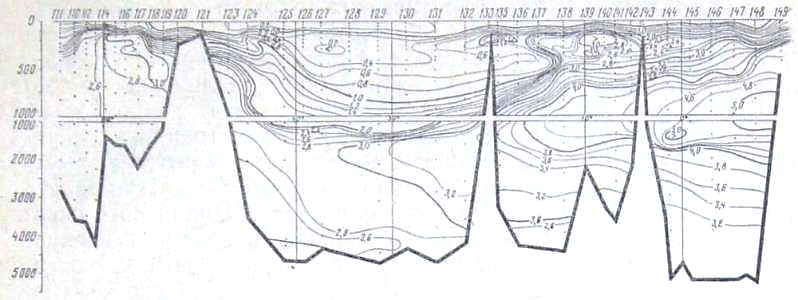
Рис. 25. Распределение дефицита кислорода на разрезе море Девиса - мыс Гвардафу
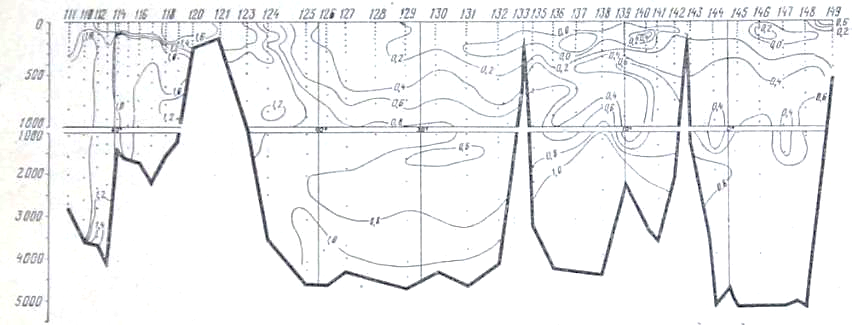
Рис. 26. Распределение преформ-фосфатов на разрезе море Девиса - мыс Гвардафуй
В глубинных водах ниже 1000 м происходит постепенное увеличение концентрации фосфатов, уменьшение концентрации кислорода и увеличение дефицита кислорода по мере движения от района Антарктики к берегам Индии. Концентрация фосфатов в слое максимума увеличивается от 2,0—2,1 мкг-ат Р/л в южных частях разрезов до 2,8—3,0 мкг-ат/л в Аравийском мере и Бенгальском заливе. Концентрация кислорода в этом же направлении уменьшается от- 350—400 мкг-ат/л в антарктических и субантарктических водах до значений менее 50 мкг-ат/л севернее 10° с. ш. Соответствующим образом ведет себя и дефицит кислорода, величины которого от 280— 320 мкг-ат/л в районе Антарктики увеличиваются до 540 мкг-ат/л (рис. 25) в северной части разреза.
Следует отметить, что уменьшение концентрации кислорода и увеличение его дефицита происходит значительно быстрее, чем увеличение концентрации фосфатов. Это легко -видеть из изложенного ниже. На всем протяжении Индийского океана горизонтальные градиенты кислорода, фосфатов и дефицита кислорода почти постоянны в глубинных водах и равны: для фосфатов 1 * Ю-7 мкг-ат Р/л/м, для кислорода 7,5 • 10-5 мкг-ат/л/м и для дефицита кислорода +5,0 * 10-5 мкг-ат/л/м. Тем не менее возрастание концентрации фосфатов и убыль содержания кислорода находятся в строгом биохимическом соответствии.
Известно (А. Редфилд, 1942), что при потреблении 1 мл кислорода высвобождается 0,373 * 10-3 мкг-ат фосфатов, или, другими словами, отношение ДР/АО, выраженное в микрограатомиой форме, при окислении органического вещества равно 420 * 10-5. В данном случае на протяжении разреза убыль кислорода, рассчитанная по уменьшению концентрации кислорода или по увеличению дефицита кислорода, равна 240 мкг-ат/л, а увеличение содержания фосфатов равно 1 мкг-ат Р/л. Вычисляя отношение ДР/ДО для глубинных вод Индийского океана, получаем величину 416 * 10-5. Как видим, эти величины (420 * 10-5 и 416 * 10-5) очень близки.
Классическое распределение кислорода и фосфатов по вертикали н Индийском океане наблюдается только ниже основного минимума кислорода. Именно на этих глубинах происходит увеличение концентрации кислорода ко дну до величин более 400 мкг-ат/л, а концентрация фосфатов достигает максимума и затем постепенно понижается с увеличением глубины.
Выше основного минимума кислорода картина распределения всех гидрохимических характеристик значительно осложняется наличием промежуточных вод различного происхождения. Построив ОР-кривые для глубинных вод ниже основного минимума кислорода (рис. 28), мы получили отрезки прямых, вытянутые вдоль «главной ОР-линии», по терминологии Постма (1964), предложенной для Тихого океана.
Дата добавления: 2022-11-28; просмотров: 2162;
Поиск по сайту
Узнать еще
- Аппарат для точечной контактной и стыковой сварки
- Выбор места для укрытия и производство земляных работ
- Генетические признаки. Деление клеток для роста. Деление половых клеток
- Демонстрации с полупроводниковыми приборами
- Изготовление станка для лучковой пилы. Уход за пилами
- Изучение вертикальной циркуляции и структуры вод арктического бассейна
- Искусственные водоёмы и уход за ними

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
