Нервная система у беспозвоночных животных
Нервную систему можно определить как организованную совокупность нервных клеток и сопутствующих им клеток иных типов (Bullock, 1977). Это определение включает в себя рецепторы, но не касается эффекторных органов, т. е. желез и мышц. У всех животных нервные клетки обладают общими чертами, по которым их легко отличить от других клеток. К таким чертам относятся градуальные электрические потенциалы в рецепторных клетках и синапсах и имеющие характер импульсов потенциалы действия, которые проводят информацию по аксонам. Эти свойства являются хотя и не универсальными, но типичными для описанных выше нейронов. Важные исключения из общего правила встречаются у некоторых примитивных животных и простейших.
Простейшие, будучи одноклеточными организмами, не имеют настоящей нервной системы. Хотя они обладают несомненными сенсорными способностями и зачаточным поведением, у них, по-видимому, нет специализированных внутриклеточных органелл, проводящих возбуждение (Bullock, 1977). Вместо этого они сами, очевидно, организованы по принципам, аналогичным тем, какие управляют физиологией нейронов. Таким образом, простейшее животное сходно с рецепторной клеткой, снабженной эффекторными органеллами.
|
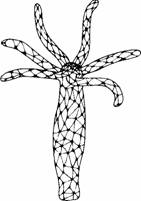
| Рис. 11.10. Нервная сеть гидры. (The Oxford Companion to Animal Behaviour. 1981.) |
Кишечнополостные - самые простые животные с истинной нервной системой. Их нервные клетки передают информацию друг другу посредством различимых нервных импульсов и синапсов. У кишечнополостных нет ЦНС, хотя иногда нервные клетки могут быть организованы в простые ганглии. Часто эти клетки образуют нервные сети, способные к диффузному проведению возбуждения по всему телу, как у гидры (рис. 11.10).
Гидра имеет рецепторные клетки, реагирующие на прикосновение и химические стимулы. У некоторых кишечнополостных встречаются также фоторецепторы. Рецепторы посылают информацию через синапсы другим нервным клеткам, но по сравнению с позвоночными нервные импульсы распространяются у них очень медленно. Нервная сеть координирует движения животного еще не совсем ясным способом. Движения совершаются медленно, и поведенческий репертуар весьма ограничен. Научение не обнаружено, если не считать формы простого привыкания. При многократных прикосновениях к ротовому отверстию гидры или актинии они сначала рефлекторно закрывают его, но при дальнейшей повторной стимуляции эта реакция постепенно исчезает. Известно, что это привыкание объясняется сенсорной адаптацией и обычно не рассматривается как истинное научение.
Среди кишечнополостных медузы обладают двумя важными особенностями более развитых нервных систем. У них имеются образования, которые, вероятно, можно было бы назвать первыми ганг-
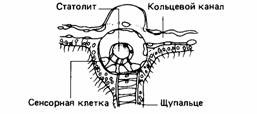
лиями и органами чувств. Эти ганглии лежат в краевых тельцах, иннервирующих фоторецепторы и статоцисты. Краевые тельца содержат четыре или более разных типов нейронов, связанных между собой (Bullock et al., 1977). Статоцисты служат для определения направления силы тяжести. В них находится маленький округлый камешек, статолит, положение которого в полости этого органа регистрируется механорецепторами в стенке статоциста, как показано на рис. 11.11. Эти рецепторы дают информацию о силе тяжести на основе распределения окружающей их не-нервной ткани, это делает статоцист настоящим органом чувств. Интересно, что такое довольно замысловатое устройство появляется одновременно с относительно сложным элементом нервной системы - ганглиями в краевых тельцах. Сочетание специализированных образований нервной системы и особых органов чувств - общая черта эволюции сенсорных процессов.
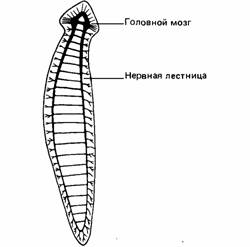
Дальнейший прогресс в организации нервной системы прослеживается у плоских червей (Plathelminthes). В отличие от кишечнополостных, но подобно большинству других беспозвоночных плоские черви обладают двусторонней симметрией, имеют голову и хвост. Рецепторы преимущественно сосредоточены в области головы, а не рассеяны по всему телу. Нервная система тоже сконцентрирована в голове в форме переднего, или головного, ганглия, который представляет собой простой мозг. От головного ганглия вдоль тела идут два нервных ствола, соединенные друг с другом нервами в виде лестницы (рис. 11.12). От нервных стволов ко всем частям тела отходят в виде сети нервные волокна. Головной ганглий и нервные стволы образуют вместе ЦНС, а сеть из нервных волокон составляет периферическую нервную систему. Такое разделение присуще большинству беспозвоночных и всем позвоночным, а у плоских червей проявляется в самой примитивной форме. Как правило, в ЦНС сосредоточена большая часть тел эффекторных нейронов, а в периферической нервной системе находятся рецепторы.
У плоских червей, например у плана-
| 
|
| Рис. 11.11.Статолит (сенсор силы тяжести) медузы. Он образован различными тканями, следовательно, является истинным органом чувств. | Рис. 11.12. Нервная система плоского червя. (The Oxford Companion to Animal Behaviour, 1981/) |
рий, сенсорные клетки на голове реагируют на прикосновение, температуру и химический состав воды. У планарий имеются также два глаза, представляющие собой скопление фоторецепторов. Сигналы от сенсорных клеток передаются в головной ганглий. По нервным стволам импульсы проводятся быстрее, чем в нервных сетях, в связи с чем поведение у плоских червей более разнообразно и определенно, чем у кишечнополостных. Они быстро обнаруживают пищу и приближаются к ней, избегают сильного света и вредных химических веществ и, по-видимому, способны к зачаточному научению. В простом Т-образном лабиринте плана-
рии научаются сворачивать предпочтительно в одну сторону, чтобы избежать прикосновения палочки. Имеются также данные о выработанных реакциях на свет, но сведения эти противоречивы.
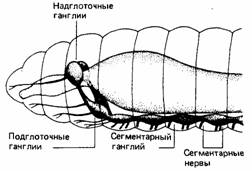
Более развитые беспозвоночные организованы по общему плану, в основе которого лежит строение аннелид, или кольчатых червей. Эти черви обладают довольно сложной нервной системой, типичная формула которой представлена у дождевых червей рода Lumbricus (рис. 11.13). В области головы находится пара надглоточных ганглиев, которые соединены с парой ганглиев, лежащих под глоткой. Эти подглоточные ганглии являются первыми в цепочке ганглиев центрального нервного ствола - по одному в каждом сегменте червя. Многие аннелиды обладают системой гигантских волокон, содержащих аксоны большого диаметра с высокой скоростью проведения, которые участвуют в реакциях быстрого избегания опасности. Рецепторы кольчатых червей большей частью представлены отдельными элементами -хемо-, механо- и фоторецепторами. У некоторых видов появляются настоящие органы чувств, например вкусовые почки в коже Lumbricus и глаза у свободноплавающей полихеты Alciope. Аннелиды обладают весьма ограниченным поведенческим репертуаром и способны лишь к зачаточному научению.
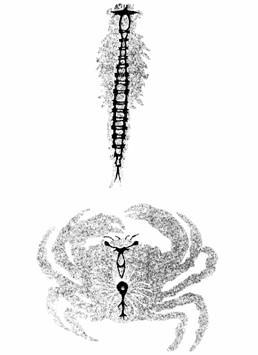
У членистоногих общий план строения нервной системы сходен с наблюдающимся у аннелид и включает центральную нервную цепочку с парой ганглиев в каждом сегменте и поперечными связями, называемыми комиссурами. У них имеется дорсальный передний мозг с окологлоточными связями, идущими к центральной цепочке. У примитивных членистоногих ясно различимо сегментарное строение, но у более развитых форм ганглии в значительной степени слиты друг с другом (рис. 11.14). Это слияние ганглиев характерно для эволюции нервной системы беспозвоночных и связано с возрастающей сложностью их сенсорной системы и поведения. Многие членистоногие, в том числе креветки, омары, скорпионы и некоторые насекомые, обладают системой гигантских волокон. Сигналы, про-
| 
|
| Рис. 11. 13. Нервная система дождевою червя. (The Oxford Companion to Annual Behaviour, 1981.) | Рис. 11.14. Цефализация уракообразных. Вверху. Гoлoвной мозг и брюшная цепочка жаброногого ракообразного. Внизу. Головной мозг и брюшная цепочка краба. |
ходящие по ним, обычно вызывают быстрый удар хвостом или прыжок, которые у этих животных составляют элемент поведения избегания.
У членистоногих больше разнообразных типов рецепторов, чем у любой дру-
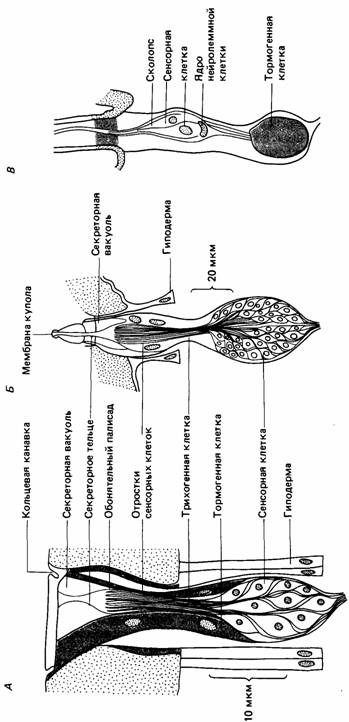
| Рис. 11.15. Примеры органов чувств у членистоногих. А. Обонятельная пластинка шершня (плакоидиая сенсилла); Б. Обонятельный конус шершня (целоконическая сенсилла); В. Волосок на антенне бабочки (трихоидная сенсилла). (По Bullock, 1977.) |
гой группы, в том числе у позвоночных (Bullock, 1977). В табл. 11.1 приведены эти основные типы. Тела сенсорных нейронов в таких рецепторах лежат близ чувствительной поверхности и не сгруппированы в сенсорные ганглии. Как показано на рис. 11.15, у одних членистоногих сенсорных нейронов мало, у других - много.
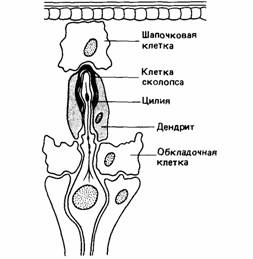
Ракообразным свойственно много типов механорецепторов, в том числе статоцисты. Особыми внутренними механорецепторами являются хордотоналъные органы, которые обычно выполняют функции проприоцепторов, отвечающих на механическое смещение (рис. 11.16). Они могут также участвовать в определении направления ветра, скорости течения воды, скорости полета, направления силы тяжести и разных типов вибраций. Механорецепторы обнаружены также в связках и мышцах, откуда они посылают информацию о положении конечностей и движении.
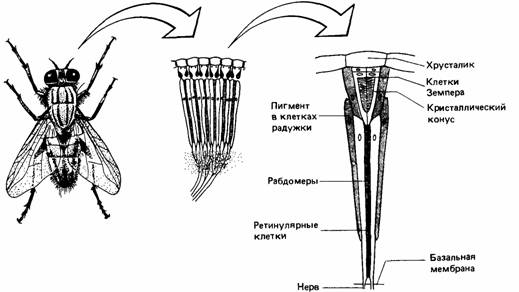
Особый интерес представляют фоторецепторы членистоногих, один из которых показан на рис. 11.17. Вообще членистоногие отличаются сложностью нервной системы и многими направлениями ее эволюционного развития, в том числе по-
|
| Рис. 11.16. Хордотональный механорецептор насекомого. (По Gray, 1950.) |
степенным переходом от сегментарного строения, свойственного кольчатым червям, к крупному мозгу, образованному многими слившимися ганглиями. Они обладают удивительно богатым поведенческим репертуаром. Некоторые виды, например медоносная пчела (подробно описанная в гл. 23), по сложности поведения
| Таблица 11.1. Некоторые типы кутикулярных рецепторов членистоногих | ||
| Структура | Морфологическое строение | Функция (известная или предполагаемая) |
| Трихоидная сенсилла | Сенсорные волоски и щетинки | Механорецепторы, проприоцепторы, звук, контактная хеморецепция, влажность, обонятельная в разных местах |
| Хетоидная сенсилла | Сенсорные шипики и щетинки | Механорецепторы, проприоцепторы |
| Лопастевидная сенсилла | Сенсорные чешуйки | Механорецепторы |
| Базиконическая сенсилла | Короткие толстые волоски; малое или большое число нейронов | Механорецепторы, контактные хеморецепторы |
| Целоконическая сенсилла | Погруженные кутикулярные конусы | Обонятельные рецепторы или рецепторы влажности |
| Ампуловидная сенсилла | Сенсорные трубочки | Обонятельные рецепторы |
| Колоколовидная сенсилла | Кутикулярные бугорки | Дирекциональные измерители натяжения |
| Плакоидная сенсилла | Кутикулярные и пористые пластинки | Неизвестна; во множестве на антеннах пчел |
|
| Рис. 11.17. Сложный глаз мухи. Каждый глаз состоит из множества омматидиев. Каждый омматидий связан с нервной клеткой, которая сигнализирует об интенсивности света в том направлении, куда обращен омматидий. Одиночный омматидий не создает образа, но, поскольку смежные омматидий обращены в слегка различных направлениях, ЦНС получает информацию о распределении интенсивности света во всем поле зрения. |
|
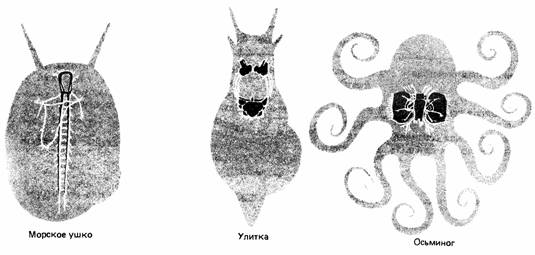
| Рис. 11.18. Примеры нервной системы моллюсков. Морское ушко (Haliotis) обладает лестничной системой. У наземной улитки (Helix) произошла значительная цефализация, а осьминог обладает самым высокоразвитым головным мозгом среди всех исследованных беспозвоночных. |
соперничают спозвоночными. Многие членистоногие способны к простому научению, хотя, по-видимому, оно связано с особыми ситуациями.
Эти эволюционные тенденции еще заметнее у моллюсков, которые достигли самого высокого уровня развития ЦНС среди всех беспозвоночных. Так. брюхоногие моллюски отличаются огромным диапазоном сложности в организации нервной системы. Например, у Haliotis (морского ушка) она сходна с системой кольчатых червей, а у сухопутной улитки произошло значительное слияние ганглиев (рис. 11.18). Наивысшего развития нервная система достигла у головоногих моллюсков. Они охотятся главным образом при помощи зрения и способны к сложному поведению, в том числе к опознанию сложных предметов и быстрому научению.
Среди головоногих, пожалуй, лучше всего изучен осьминог. Его головной мозг содержит около 170 миллионов нервных клеток (у крупных ракообразных их примерно 100 000) и состоит приблизительно из 30 разных долей, многие из которых обладают собственными функциями (рис. 11.18). Больше половины нервной ткани мозга составляют зрительные доли. Они соединены с парой крупных глаз, наиболее развитых среди беспозвоночных и соперничающих с глазами позвоночных (см. гл. 12).
По сложности поведения между моллюсками имеются большие различия, но наиболее продвинуты с этой точки зрения головоногие. Эта группа тщательно изучена исследователями поведения (например, Wells, 1966) в связи с поразительной способностью к научению.
Дата добавления: 2016-06-18; просмотров: 5834;
Поиск по сайту
Узнать еще
- Arthropoda. Клещи. Систематика. Морфология. Медицинское значение.
- Arthropoda. Паукообразные. Систематика. Географическое распространение. Морфология. Скорпионы. Пауки. Медицинское значение.
- Arthropoda..Систематика.Насекомые.Морфология.Классификация.Медицинское значение.
- Arthropoda.Систематика.Блохи.Виды блох.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское и эпидемиологическое значение.Меры борьбы.
- Arthropoda.Систематика.Мошки,мокрецы,слепни,оводы.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское значение,меры борьбы.
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- D) Система класифікації за підтримкою багато гілкового виконання програми.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
