Наследование поведенческих признаков
«Вопрос о том, что важнее в формировании физических черт человека и его личности - природа или воспитание, генотип или окружающая среда, - попросту ошибочен и может ввести в заблуждение. Генотип и окружающая среда важны в равной степени, так как и то и другое необходимо... Вопрос о роли генотипа и окружающей среды в развитии человека следует поставить так: до какой степени различия, наблюдаемые между людьми, определяются различиями их генотипов и различиями условий, в которых люди рождаются, растут и воспитываются? (Dobzhansky, 1964).»
Большая часть работ Фрэнсиса Гальтонаоснована на представлении о том, что фенотипическое сходство между родственниками частично определяется сходством их генотипов. Однако Гальтон не был знаком с работами Менделя и считал, что наследственность носит слитный характер. Он думал, что материал наследственности непрерывен, поэтому признаки потомков промежуточны между признаками их родителей. На основе таких допущений Гальтон впервые разработал методику расчета корреляций между родственниками как средство оценки наследования тех или иных признаков. В 1918 г. Фишер опубликовал статью, в которой было показано, что возможно предсказать корреляции, ожидаемые между родственниками на основе теории Менделя. Работы Менделя стали известны общественности в 1900 г., однако они относились только к дискретным признакам. Фишер попытался применить менделевскую теорию дляанализа непрерывных признаков, таких, как рост и масса. Интересно, что его выводы были сходны с выводами Гальтона. причем большая часть прогнозов в этих двух теориях была почти идентична.
Наследуемость впервые была определена как доля изменчивости, связанная с различиями в наследственности (Lush, 1940). Иными словами, когда мы анализируем конкретный признак в популяции животных, такой, например, как масса тела, мы определяем, какой вклад вносят в фенотипическое проявление этого признака генотипическая и средовая компоненты. Рассматривая родственные популяции в различных условиях, иногда можно оценить степень влияния на изменчивость особей факторов окружающей среды и генетических факторов.
Существуют разнообразные статистические методы оценки наследуемости поведения. Некоторые основаны на исследовании фенотипической изменчивости у генотипически однородных особей. Эта общая фенотипическая изменчивость генотипически идентичных особей сравнивается с общей фенотипической изменчивостью в природной генотипически разнообразной популяции. Такое сравнение дает соотношение, в котором противопоставляется генотипическая компонента фенотипического разнообразия средовой компоненте. При оценке с помощью этой статистической методики мы сталкиваемся с определенными трудностями, но она ясно показывает, насколько ошибочно относить один вид поведения к «наследственному», а другой - к «приобретенному». В случае если возникнет новая генная вариация или популяция попадает в другие условия среды, это соотношение может сильно измениться. Типы поведения, которые «генетически детерминированы» в одной среде (у монозиготных близнецов, воспитанных вместе, фенотипы оказываются сходными), могут быть подвержены и сильному влиянию средовых факторов, например климатических (у монозиготных близнецов, воспитанных порознь, фенотипы оказываются разными). Напротив, тип поведения, который «детерминирован средовыми факторами» (у всех генотипически различающихся особей одинаковый фенотип, если их воспитывают в одинаковых условиях), может, очевидно, в значительной степени подвергаться воздействию генетической компо-
| Рис. 4.11. Среднее число ошибок, сделанных в лабиринте «умными» и «глупыми» крысами, выросшими в обогащенной, нормальной и обедненной среде. (По Cooper, Zubek, 1958.) | 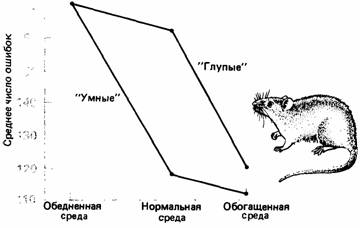
|
ненты в среде, в которой проявляется генотипическое разнообразие. Все эти взаимодействия позволяют лишь в строгих пределах определить степень наследуемости того или иного типа поведения, а также провести границу между врожденным и приобретенным поведением. Мы можем сказать, что данный тип поведения может наследоваться в 80% случаев при данном генотипе, в данной среде и в конкретной популяции, но мы не можем переносить эти выводы на другую популяцию или среду.
При оценке наследуемости ошибки могут проистекать из разных источников, наиболее важный из них - это взаимодействие генотипа и среды и корреляции генотипа и среды. Взаимодействие между генотипом и средой вводит изменчивость, которая не принимается во внимание при обычных расчетах наследуемости. Например, Трайон (Тгуоп, 1942) проводил селекцию крыс на неспособность ориентироваться в лабиринте («глупые» крысы) и на блестящее ориентирование в нем («умные» крысы). Этих крыс выводили и тестировали в обычных лабораторных условиях. А Купер и Зубек (Cooper, Zubek, 1958) выращивали одних крыс в обычных условиях, других - в обедненных, без укрытий и препятствий, а третьих - в условиях, обогащенных сложными лабиринтообразными сооружениями. При испытаниях в стандартном лабиринте эти две линии крыс, выращенные в обычных условиях, сильно различались по своим возможностям ориентироваться в лабиринте, как это обнаружили раньше и другие исследователи. Однако обе линии крыс, выращенных в обедненных условиях, одинаково плохо ориентировались в лабиринте, тогда как обе линии, выращенные в обогащенной среде, ориентировались одинаково хорошо (рис. 4.11). Хотя эти две линии отчетливо различаются генотипически, в фенотипе эта разница проявляется только у крыс, выращенных в определенных условиях среды.
Корреляция генотипа и среды - другой потенциальный источник ошибок при оценке наследуемости поведенческих признаков - может возрастать, когда особи избирают какую-то особую среду или у них развиваются определенные привычки для компенсации генотипических дефектов. Однако, несмотря на эти трудности, оценка наследуемости оказалась особенно полезной для разделения эффектов природы и воспитания.
Оценки наследуемости, основанные на сходстве между родственниками, часто использовались в генетическом анализе поведения человека. Однако здесь есть ряд осложняющих факторов. Во-первых, генетическое доминирование снижает корреляцию между родственниками. При генетическом доминировании вклады родителей в проявление признака неодинаковы, т. е. вводится еще один источник изменчивости. Это называется дисперсией доминирования. Однако дисперсия доми-
нирования оказывает различное действие на корреляции родителей и потомков и корреляции между сибсами. Фактически разница между этими двумя коэффициентами корреляции должна точно составлять одну четверть дисперсии, обусловленной доминированием (Bodmer, Cavalli-Sforza, 1976).
Во-вторых, подбор пар у человека, вероятно, не является случайным в отношении некоторых признаков. Можно допустить, что в отношении определенной части признаков, которые нельзя наблюдать непосредственно, например уровней ферментов, подбор пар происходит случайным образом, однако этого нельзя сказать в отношении таких признаков, как рост и коэффициент умственного развития. Корреляции между мужем и женой по этим признакам обычно положительны, что указывает на неслучайный или выборочный подбор, поскольку люди обычно стремятся выбирать партнеров одинакового роста и умственного развития. Эти корреляции по росту составляют примерно +0,3, а по умственному развитию - примерно + 0,4. На эти корреляции могут влиять и социальные факторы, так как между социально-экономическим положением и данными признаками также существует положительная корреляция. Поэтому тенденция выбирать мужа или жену из той же социальной группы обычно приводит к положительной корреляции между мужем и женой. Ассортативные браки у человека могут влиять на увеличение частоты гомозигот.
В-третьих, при анализе наследуемости поведения человека необходимо правильно оценивать средовую дисперсию, поскольку в этом случае условия среды нельзя контролировать, как при исследованиях, проводимых на животных. Был предложен способ (Cavalli-Sforza, Bodmer, 1971) разделения средовой дисперсии на отдельные компоненты. Прежде всего следует различать внутрисемейную дисперсию. Она может быть обусловлена возрастными различиями, порядком рождения, различиями по полу и т.д. В семьях разных размеров эта дисперсия может также зависеть от уровня питания и наличия жилищной площади.
Далее существует дисперсия, обусловленная различиями между семьями одного социально-экономического слоя. Некоторую информацию о значении этого фактора можно почерпнуть из рассмотрения корреляции между приемными детьми и их приемными родителями, однако результаты могут быть тенденциозны из-за того, что родителей выбирают посреднические агентства.
Затем следует учитывать дисперсию между социально-экономическими слоями. Культурная преемственность может привести к корреляциям между родственниками, которые трудно отличить от корреляций, обусловленных генотипическими факторами. Культурная преемственность между родителями и детьми в значительной степени маскирует биологическую наследуемость (Cavalli-Sforza, Feldman, 1974). Это остается одной из самых интересных проблем в биологии.
Возможно, следовало бы также учитывать средовую дисперсию, сопровождающую расовые различия. Например, условия жизни черного и белого населения США сильно различаются. Это может быть обусловлено частично культурными различиями, частично социально-экономическими факторами.
И наконец, существует дисперсия, обусловленная взаимодействием генотипа и среды. У человека такое взаимодействие может иметь место, например, между возрастом наступления половой зрелости и типом школьного обучения, доступным разным возрастным группам.
В общем, мы можем конкретизировать средовую дисперсию, добавляя компоненты, перечисленные выше.
Кавалли-Сфорца и Бодмер (Cavalli-Sforza, Bodmer, 1971) пришли к выводу, что оценки наследуемости не соответствуют относительному значению природы и воспитания; исключения составляют некоторые конкретные обстоятельства. Изменения в условиях среды могут свести на нет даже прямые измерения наследуемости. Например, для особенностей телосложения человека характерна высокая степень наследуемости, однако в популяциях человека эти особенности будут наследоваться по-разному, что обусловлено раз-
личными факторами среды, например питанием. Если оценка наследуемости позволяет нам судить об отношении преобладающих генетических различий к преобладающим различиям среды, то эти оценки нельзя относить к другим популяциям, другой среде или другому периоду времени.
ДЛЯ ЗАПОМИНАНИЯ
1. В этой главе мы рассмотрели некоторые методы исследования генотипического влияния на поведение животных. Мы увидели, что в некоторых случаях можно определить действие одного гена, однако обычно даже простые аспекты поведения контролируются множеством генов.
2. Полигенное наследование можно изучать в экспериментах по скрещиванию, спланированных так, чтобы обнаружить влияние хромосомных мутаций на поведение или чтобы обнаружить количественные различия в поведении различных генетических линий.
3. Наследуемость поведенческих признаков оценивается дисперсией фенотипических признаков в конкретной популяции. Наследуемость признака - это часть наблюдаемой дисперсии, обусловленной различиями в наследственности. Наследуемость можно изучить, рассматривая различия между близкородственными животными, выросшими в различных условиях, и различия между животными, выросшими в одинаковых условиях.
Рекомендуемая литература
Bodmer W.F., Cavalli-Sforza L. L. (1976) Genetics, Evolution, and
Man, Freeman, New York.
Ehrman L,, Parsons P. A. (1976) The Genetics of Behavior, Sinauer
Associates, Sunderland, Massachusetts.
Дата добавления: 2016-06-18; просмотров: 2890;
Поиск по сайту
Узнать еще
- I.5. Наследование группы крови
- II.6. Наследование резус-принадлежности
- АНАЛИЗ ВЗАИМОСВЯЗЕЙ КАЧЕСТВЕННЫХ ПРИЗНАКОВ
- Аутосомно-доминантное наследование
- В развитии признаков пола
- Виды признаков и шкал
- ВОПРОС 1. Сцепленное наследование генов.
- Вопрос № 22. Коэффициенты оценки связи качественных признаков, представленных двумя градациями. Коэффициент ассоциации и контингенции.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
