ШИРОКОЛИСТВЕННЫЕ И СМЕШАННЫЕ ЛЕСА
УМЕРЕННЫХ ШИРОТ
Географическое распространение,
гигротермический и геохимический режимы
Широколиственные летнезеленые леса, иногда с большой примесью хвойных пород, распространены в условиях достаточного в течение всего года увлажнения и периодического снижения температур зимой, приводящего к полной остановке вегетации древесного яруса. Широколиственные летнезеленые леса субсредиземноморского типа с подъемом температур в середине лета получают меньше осадков. В широколиственных летнезеленых лесах более северных широт одновременно с подъемом температур обычно наблюдается и увеличение количества осадков. В зависимости от степени континентальности зимы могут быть почти безморозными и бесснежными (Колхида, некоторые районы Приатлантической Европы) или же характеризуются устойчивыми морозами и снежным покровом (Южная Россия). Различные сочетания климатических факторов в пределах ареала летнезеленых лиственных лесов показаны на приведенных климадиаграммах (рис. 35).
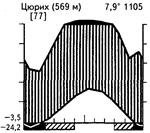
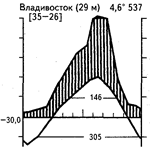
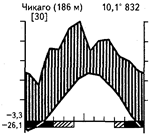
Рис. 35. Климадиаграммы различных районов с экосистемами широколиственных и смешанных лесов умеренных широт
В южном полушарии изолированный массив летнезеленых лесов существует лишь в Чили и в прилегающих районах Аргентины на широте 38-40°, а южнее снова идут вечнозеленые леса умеренных широт. Южноамериканский массив летнезеленых лесов образован одним видом южного бука-нотофагуса (Nothofagus obliqua), местное название которого «робле», что по-испански означает «дуб». Действительно, леса из робле внешне очень похожи на дубняки из пушистого дуба (Q. pubescens), характерные для субсредиземноморских формаций юга Европы. По режиму тепла и влаги они также сходны.
В дальнейшем речь пойдет только о широколиственных летнезеленых (неморальных) лесах северного полушария, которые распространены в Европе, на Дальнем Востоке и небольшими изолированными пятнами в горах Евразии, в Северной Америке, особенно на востоке этого материка, в районе Аппалачских гор. Неморальные формации Восточной Азии и Аппалачей к югу постепенно переходят в субтропические вечнозеленые леса.
Под неморальными формациями образуются бурые и серые лесные почвы. Первый тип свойствен территориям с очень мягкой зимой и влажным прохладным летом, он связан, в частности, с буковыми лесами. Второй тип приурочен к более континентальным районам с теплым и сухим летом; часто серые лесные почвы формируются под дубравами из черешчатого дуба (Q. robur) в лесостепной зоне. В бурых лесных почвах происходит энергичное образование вторичных глинных минералов, вынос карбонатов. Однако выраженного подзолообразовательного процесса часто не происходит, что связывают с обилием опада, обогащенного кальцием. Реакция в почвенном профиле обычно слабокислая и гидраты полуторных окислов Fe и А1 малоподвижны; Содержание гумуса обычно равно 3-5%.
Серые лесные почвы имеют промывной режим только весной, во время снеготаяния. Это приводит к глубокому выносу веществ в мощные иллювиальные горизонты. До глубины 50-80 см почва имеет кислую реакцию, но на глубине 150-200 см формируется карбонатный горизонт. Богатый кальцием опад, как и в бурых почвах, оказывает существенное влияние на почвенные процессы. В верхнем горизонте содержание гумуса может достигать 9%, но обычно оно равно 4-5%.
В наиболее теплых и одновременно влажных районах летнезеленых лесов (например, в Колхиде и в Ленкорани) происходит разложение не только первичных, но и вторичных минералов с образованием каолина и свободного кремнезема, глинозема и окислов железа. Таким образом, имеются уже черты аллитизации почвенной толщи, а также своеобразные черты оподзоливания из-за кислой реакции и выраженного промывного режима. Такого рода почвы очень характерны также для вечнозеленых муссонных влажных субтропических лесов юга Восточной Азии, Юго-Восточной Австралии, гемигилеи Чили и для некоторых горных тропических лесов. Красноземы и желтоземы отличаются от бурых лесных почв более интенсивным химическим выветриванием. Нередко широколиственные леса с опадающей зимой листвой и вечнозеленые влажные субтропические леса образуют плавный переход, в котором изменение типа почвы наступает раньше, чем смена листопадных пород вечнозелеными.
Сообщества фотоавтотрофов
Летнезеленые широколиственные леса обычно образованы небольшим (1-3) числом доминирующих видов деревьев, и только самые богатые типы их в Восточной Азии и в Аппалачах имеют полидоминантные древостой с 10-20 и более видами. Обычно хорошо развит верхний сомкнутый ярус, второй древесный ярус более разрежен и может вообще отсутствовать. Высота спелого леса обычно близка к 30 м. Кустарниковый ярус сильно изменчив — от почти сплошного до крайне разреженного. Это же касается и травянистого покрова, который образован многолетними мезофильными растениями - гемикриптофитами и геофитами. Последние в значительной степени образованы эфемероидами, которые развиваются и цветут ранней весной, когда до распускания листьев на деревьях под пологом сохраняется высокая освещенность. После распускания листвы освещенность резко падает - к почве проходит лишь 1-2% светового потока. Часть травянистых растений и кустарников состоит из вечнозеленых растений, сохраняющих зимой листья под снегом. В наиболее мягких условиях некоторые растения под пологом леса могут цвести уже в конце зимы (например, цикламены). Характерно полное отсутствие однолетних трав.
Среднеевропейские неморальные сообщества отличаются наиболее бедным флористическим составом, что связано с недавними оледенениями. В западной части наиболее обычно доминирование бука (Fagus sylvatica) и в меньшей степени - скального дуба (Quercus petraea). Чуть далее к востоку, включая западные области Украины часто доминирует граб (Carpinus betulus). В восточной половине Европы основной породой широколиственного леса становится дуб черешчатый, часто вместе с липой (Tilia cordata). В лиственных лесах постоянно встречаются также клены (Acer platanoides, A. pseudoplatanus), ясень (Fraxinus excelsior), вязы (Ulmus scabra, U. laevis). В наиболее теплых районах субсредиземноморского типа листопадные леса образованы дубом пушистым (Q. pubescens) и несколькими близкими видами (Q. pyrenaica, более восточным Q. cerris). В Закавказье обычны каштанолистный дуб, грузинский дуб (Q. castaneifolia, Q. iberica) и еще несколько видов. Отдельными сравнительно небольшими участками на западе и юге ареала неморальной растительности Европы и Закавказья встречаются каштан, орех (Castanea, Juglans), а также многие другие древесные породы -реликты плиоценовой неморальной растительности Европы, в том числе один из самых массовых эдификаторных видов гирканских листопадных лесов - железное дерево (Parrotia persica), образующее леса в Талыше и в горах Эльбурс. В горных лиственных лесах Кавказа, кроме упомянутых видов дубов, растет и бук, часто образующий чистые древостой. Кавказский бук очень близок к европейскому, но все же выделяется в особый вид Fagus orientalis.
На западе Европы и на Кавказе в подлеске лиственных лесов часто разрастается вечнозеленый падуб (Ilex aquifolium).
На Кавказе к нему добавляются и вечнозеленые рододендроны. Ареал падуба хорошо соответствует нулевой январской изотерме. Особенно типичны листопадные кустарники подлеска, из которых наиболее распространены лещина (Corylus avellana), различные виды жимолости (Lonicera), ивы, рябины, бересклета, калины и др. В поймах небольших речек обычно растут ольха (черная или серая) и различные ивы с примесью других пород. Лианы и эпифиты обычно малочисленны. Из лиан чаще встречаются различные виды ломоносов (Clematis) и плющ (Неdera helix) - представитель семейства аралиевых с вечнозелеными листьями. Эпифиты включают мхи и лишайники. Лишь в особо благоприятных условиях (влажные леса с мягкими зимами) становятся заметными и некоторые эпифиты из высших растений (обычно папоротники).
Отдельные массивы неморальных лесов распространены в горах Средней Азии и далее на юго-восток. Они занимают очень ограниченные площади, хотя крайне интересны по своему составу. Таковы, например, орехово-плодовые леса Ферганского и Чаткальского хребтов Западного Тянь-Шаня с господством ореха (Juglans regia), но включающие в себя разнообразные виды кленов, яблонь, алычи; сходные сообщества (чернолесье) представлены также в Гиссаро-Дарвазе.
Флористически наиболее разнообразны неморальные сообщества Восточной Азии, представленные и у нас в стране на юге Приморья. Многие эдификаторные виды европейских лиственных лесов заменяются здесь другими видами того же рода. Например, вместо европейского и кавказского буков на Дальнем Востоке представлены бук Зиболда, японский, Энглера, длинночерешковый (Fagus sieboldii, F. japonica, F. engleriana, F. longipetiolata). Первые два вида обитают на островах Японии, вторые два - на материке, южнее наших границ. Листопадные дубы также образуют особые виды на востоке Евразии: дуб монгольский (Q. mongolica - одна из главных лесообразующих пород, однако именно в Монголии не встречается), дуб зубчатый (Q. dentata - в пределах нашей страны встречается лишь на южных Курилах, обычен в Японии, Корее, Китае). Есть и специфические ясени (F. mandshurica, F. rhinchophylla).
Дальневосточные широколиственные и хвойно-широколиственные леса отличаются разнообразием древесных видов. В Приморье, кроме преобладающего дуба монгольского, обычны также липа маньчжурская (Tilia mandschurica), клен мелколистный (Acer mono), диморфант (Kalopanax ricinifolia), амурский бархат (Phellodendron amurense), мелкоплодник ольхолистный (Micromelis ainifolia). В приречных сообществах появляются также тополь корейский (Populus koreana), чозения (Chosenia macrolepis), клен маньчжурский (A. mandschuricum) и маньчжурский орех (Juglans mandschurica). Кроме лиственных пород деревьев, встречаются и хвойные. Особенно характерна пихта цельнолистная (Abies holophylla), часто встречается также корейская кедровая сосна, называемая кедром (Pinus koraiensis), изредка попадается тисе остроконечный (Taxus cuspidata).
Обычно хорошо развит также второй ярус древостоя из граба сердцелистного (Carpinus cordata), низкорослых кленов (A. pseudosieboldianum, A. ukurundense), черемухи Маака (Padus maackii), вишни Максимовича (Cerasus maximoviczii), маакии (Маackia amurensis), сирени амурской (Syringa amurensis).
В подлеске обитают разнообразные кустарники, в первую очередь лещина (Corylus heterophylla), элеутерококк (Eleutherococcus senticosus), акантопанакс (Acanthopanax sessiliflorum), аралия маньчжурская (Aralia mandschurica), разные виды жимолости, бересклета, смородины. Обращают на себя внимание и крупные лианы - виноград (Vitis amurensis, V. thunbergii), лимонник (Schizandra chinensis), актинидия (Actinidia kolomikta) и кирказон маньчжурский (Aristolochia mandschuriensis).
В травяном покрове встречаются некоторые виды, общие или близкие с европейскими широколиственными лесами, например майник двулистный (Majanthemum bifolium). В то же время бросаются в глаза виды, экзотические с точки зрения европейца, часто представляющие особые роды и даже семейства, отсутствующие в летнезеленых лесах Европы.
В американском неморальном секторе леса состоят из многочисленных особых видов дуба, бука, ясеня, клена, липы, каштана, вяза, ореха, березы, тополя, гикори (Carya), ниссы (Nyssa), ликвидамбара (Liquidambar), тюльпанового дерева (Liriodendron tulipifera), сассафраса (Sassafras officina1е), платанов, белой акации (Robinia pseudoacacia), гледичии (Gleditschia triacanthos). Два последних вида широко распространены у нас в культуре на юге Крыма и Кавказа, на Украине, а местами и севернее. Столь же богаты подлесок и второй ярус древостоя. Характерны также лианы из различных видов Виноградовых (Vitis, Ampelopsis, Partenocissus) и других семейств, флористически район вокруг Аппалачей весьма сходен с Восточной Азией. Здесь множество общих родов и викарирующих видов, свидетельствующих о былых взаимосвязях. По числу сохранившихся примитивных семейств и родов Восточная Азия превосходит Северную Америку, однако богатство конкретных фитоценозов весьма варьирует, и часто именно американские леса оказываются богаче из-за полидоминантного характера не только травяного яруса, но даже древостоя, который в отдельных местах может содержать несколько десятков видов.
Наиболее обычные группировки в Аппалачах состоят из различных видов дуба (Quercus borealis, Q. alba, Q. falcata, Q. prinus, Q. minor, Q. marilandica, Q. lyrata, Q. nigra) с участием двух видов ореха, каштана (Castanea dentata), платана, кленов. Каштаны сейчас сильно пострадали от ввезенного из Китая паразитического гриба. Обычны также буковые леса (F. grandiflorus) с сахарным кленом (A. saccharum), американской липой (Т. americana), желтой березой (Betula lutea), тюльпановым деревом. Западнее Аппалачей среди основных лесо-образующих пород часто можно видеть также гикори многих видов, чаще всего Carya pecan. Виды гикори очень близки к настоящим орехам (Juglans). Гикори образуют смешанные древостой с дубами, ясенями, кленами и другими породами. Пояс гикори сильно пострадал от пожаров и рубок. В южной части неморального массива Атлантической Америки становятся заметными вечнозеленые кустарники и низкие деревья, например персея бурбонская (Persea burbonia) из лавровых, магнолия крупноцветная и др. На юге обычными становятся также сосняки из теплолюбивых видов сосны с примесью дубов, ликвидамбара и других листопадных пород.
К северу, в Канаде, видовое разнообразие лиственных лесов падает. Большая часть их образована сахарным кленом, американской липой, желтой березой, белым дубом, американским ясенем. Здесь же обычны смешанные леса с добавлением из хвойных - нескольких сосен, елей, туи восточной и канадской тсуги. В Северной Америке обычная широтная протяженность полосы широколиственных лесов заменяется почти меридиональной, поэтому отдельные неморальные элементы проникают далеко к северу, почти до границ лесотундры. В Евразии сходное явление в более ограниченных масштабах можно проследить лишь в крайних западных и восточных океанических секторах материка (например, в Шотландии и на Камчатке). Множество реликтовых местонахождений неморальных элементов свидетельствует об их былом очень широком распространении. Это подтверждается и палеогеографическими данными, которые показывают преобладание широколиственных лесов на огромных пространствах северного полушария во время третичного периода (тургайская флора). Впрочем, в их составе большую роль играли и многие хвойные породы, сохранившиеся на родовом уровне сейчас главным образом в «убежищах» Восточной Азии и Северной Америки (секвойи, метасеквойи, тсуги и т. п.).
Гетеротрофы первого порядка
Мощная почвенная толща, значительные запасы гумуса и большая глубина гумусового горизонта, хорошо развитая подстилка - все это благоприятствует существованию богатой почвенной фауны, значительная часть которой представлена гетеротрофами первого порядка, потребителями опада на разной стадии его разложения. Активность почвенных микроорганизмов, макромицетов и животных в зимний период резко падает. Но некоторые группы остаются деятельными даже под снегом, который обычно защищает почву от промерзания.
Первое место по запасам биомассы среди гетеротрофов первого порядка принадлежит дождевым червям-люмбрицидам, хотя по величине чистой вторичной продуктивности почвенные микроорганизмы (бактерии, грибы, актиномицеты) оказываются впереди. Для лиственного леса Европы общая зоомасса почвенных животных составляет 1 т, а почвенных микроорганизмов - 0,3 т на 1 га, т. е. соответственно 100 и 30 г/м2. При этом на долю дождевых червей приходится 60 г/м2. В конкретных биоценозах указанные цифры могут значительно колебаться (чаще в меньшую сторону).
Важное место среди детритофагов занимают разнообразные личинки насекомых, а также комплекс микроартропод, в первую очередь панцирные клещи (Oribatei) и ногохвостки (Collembola). Последние представлены в каждом конкретном месте комплексом из 15-20 видов, из которых более половины особей приходится на 3-4 вида. Эти доминирующие виды, как правило, принадлежат к родам Onychiurus, Folsomia, Isotoma. Общая численность микроартропод-детритофагов нередко превышает 100 тыс. экз/м2, из которых обычно не менее половины приходится на ювенильные стадии. Клещи и коллемболы составляют не менее 80% (чаще более 90%) от всех микроартропод. Суммарная биомасса (живая) этих животных в лиственных лесах колеблется в широких пределах, чаще она близка к 1 г/м2. Общий запас зоомассы в пределах 1-2 г/м2 характерен также для кивсяков, нематод (при численности в несколько десятков миллионов экземпляров на 1 м2), энхитреид. В наиболее влажных и теплых сообществах особенно сильно увеличивается численность кивсяков, их масса при этом может превысить 10 г/м2.
Весьма типичны для многих лиственных лесов следующие средние показатели живой массы почвенных беспозвоночных (в г/м2): сапрофаги - 97,4, в том числе дождевые черви - 91,2; нематоды - 1,5; кивсяки - 2,1; энхитреиды - 1,6; коллемболы (ногохвостки) - 0,4; панцирные клещи - 0,2; прочие клещи -0,1; фитофаги - 2,0; хищники и паразиты - 0,7. В почвенном ярусе сосредоточено более 90% общей зоомассы, здесь представлены практически все сапротрофы.
Фитофаги (хлорофитофаги) представлены как в почве, так и в верхних ярусах. Напочвенные беспозвоночные имеют примерно такую же зоомассу, как и почвенные. Потребляющие растительные корма позвоночные животные (грызуны, копытные) лишь немного уступают напочвенным беспозвоночным-фитофагам. Так, в дубравах под Курском биомасса почвенных беспозвоночных-фитофагов составляла 2,0 г/м2, напочвенных беспозвоночных фитофагов - 1,5 г/м2, почвенных позвоночных-фитофагов - 0,01 г/м2, напочвенных позвоночных-фитофагов - 1,0 г/м2. Лишь в годы массовых размножений зеленой дубовой листовертки (Tortrix viridana) зоомасса напочвенных беспозвоночных-фитофагов может возрастать до 15 г/м2.
На рисунке 36 приведена схема распределения на дубе 19 видов его наиболее обычных потребителей. Кроме указанных беспозвоночных, в европейских лесах характерны такие позвоночные-хлорофитофаги, как благородный олень (иногда лось), косуля, рыжая полевка (Clethrionomys glareolus), лесная и желтогорлая мыши (Apodemus sylvaticus, A. flavicollis). По существу всеядный кабан все же в большей степени питается растительными кормами. В лиственных лесах, как и в лесах других типов, большая часть хлорофитофагов существует за счет зеленой массы - листьев и побегов растений. Однако весьма важна также группа потребителей семян (главным образом древесных растений - желудей, семян бука, липы и др.), к которой в значительной степени принадлежат упомянутые лесная и желтогорлая мыши. Хлорофитофаги-позвоночные представлены почти исключительно млекопитающими.

Рис. 36. Потребители дуба (по П. Дювиньо и М. Тангу, 1968)
Листья: 1 — долгоносик-прыгун (Orchestes quercus); 2 — трубковерт (Attelabus);
3 — майский хрущ (Melolontha melolontha); 4— златогузка (Euproctis chrysorrhoea);
5 — кольчатый шелкопряд (Malacosoma neustria); 6 — пяденица-обдирало (Hibernia defoliaria); 7 — зимняя пяденица (Cheimatobia brumata); 8 — зеленая дубовая листовертка (Tortrix viridana).
Желуди: 9 — желудевый долгоносик (Balaninus glandium).
Почки: 10 — грушевый листовой слоник (Phyllobius piri).
Ветви: 11 — темная мягкотелка (Cantharis obscura).
Кора: 12 — зеленая узкозлатка (Agrilus viridis); 13 — дубовый заболонник (Scolytus intricatus); 14 — дровосек-рагий (Rhagium).
Древесина: 15 — большой дубовый усач (Cerambyx cerdo); 16 — жук-олень (Lucanus cervus).
Корни: 17 — майский хрущ, личинка; 18 — хлебный полосатый щелкун (Agriotes linatus); 19 — корневая орехотворка (Biorrhiza aptera)
Доля птиц в общей массе этой группы крайне мала и не превышает обычно нескольких процентов (чаще меньше 1%).
Комплекс хлорофитофагов из позвоночных, как и из беспозвоночных, существенно меняется в разных регионах. Так, например, в Северной Америке, к общим видам копытных, как, например, лось и благородный олень, называемый здесь вапити, прибавляется мелкий белохвостый олень, во многом аналогичный косуле. Очень резко различен комплекс грызунов, так как представители семейства мышиных (Muridae) в Новый Свет не проникают (лишь в историческое время человек завез туда домовых мышей и крыс). Вместо лесной и желтогорлой мышей здесь обитает весьма сходный с ними по экологии представитель семейства хомякообразных (Cricetidae) белоногая «мышь», или белоногий хомячок (Peromyscus leucopus). Обычны еще некоторые виды грызунов, одни из общих с Евразией родов (например, Microtus ochrogaster, M. pinetorum, Sciurus niger, Marmota monax), другие же из экзотических для европейца родов (Synaptomys cooperi, Ochrotomys, Reithrodontomys, Sylvilagus floridanus, последний вид - из зайцеобразных). Для Америки очень типично большое разнообразие бурундуков (по данным J. Whitaker, 1997 - 22 вида, в то время как в Евразии - только один таежный вид), некоторые виды живут в лиственных лесах. К ним примыкают также выделяемые в особый род бурундуковые, или красные, белки (Tamiasciurus - 2 вида). Наконец, следует упомянуть также не имеющего экологических эквивалентов в Старом Свете древесного дикобраза-поркупина (Erethizon dorsatus) из особого семейства, свойственного Новому Свету. Поркупин широко населяет не только широколиственные, но и другие леса от Канады до Мексики. Широко распространен и всеядный опоссум (Didelphis marsupialis) -единственный представитель отряда сумчатых, проникший из Южной Америки.
Гетеротрофы высших порядков
Внутри каждого размерного блока сообщества можно указать характерные группы гетеротрофов высших порядков, живущих за счет сапрофагов и хлорофитофагов. Так, в мире почвенных микроорганизмов большинство простейших (инфузории, корненожки, некоторые жгутиковые) питается бактериями. Среди наиболее мелких многоклеточных животных - нематод, коловраток, тихоходок - также имеются хищники, которые поедают микроорганизмы и представителей своей же группы. Преимущественно сапрофаговые панцирные клещи и ногохвостки все же, вероятно, в значительной степени относятся и к высшим трофическим уровням, так как существенное место в их питании принадлежит грибам. Но среди микроартропод есть и выраженные хищники, численность которых обычно в сотни раз уступает двум доминирующим группам.
Это мельчайшие двухвостки-диплюры, ложноскорпионы, свободноживущие гамазовые клещи. Наиболее крупных беспозвоночных поедают такие хищники, как многоножки-геофилы и костянки, жуки-стафилины и жужелицы (хищничают и личинки, и взрослые стадии). Наконец, исключительно разнообразные по видовому составу пауки также представляют гетеротрофов высшего порядка среди беспозвоночных.
Кроме хищников, к этой группе относятся и уже неоднократно упоминавшиеся паразитоиды - наездники-ихневмониды, бракониды, хальциды, некоторые другие перепончатокрылые, а также и двукрылые (мухи-тахины, например).
Как во всех ранее рассмотренных сообществах, в неморальных биоценозах важнейшей группой хищников являются муравьи. В первую очередь это относится к крупным видам рода Formica, близким к евразиатским Formica rufa. Впрочем, эта группа столь же типична и для бореальных лесов северного полушария.
Участие хищных и паразитических животных в создании общего фонда зоомассы ничтожно и обычно много меньше 1 %. Однако регуляторные функции их совершенно необходимы для нормального существования экосистемы.
Как и в других группах организмов, разные части ареала неморальных комплексов имеют существенно различные видовые составы хищных форм. Это можно видеть на примере некоторых групп позвоночных животных.
Представленные в широколиственных лесах северного полушария амфибии и рептилии практически полностью существуют за счет хищничества (исключая головастиков амфибий). В Европе в число этих животных входят, например, травяная и остромордая лягушки, серая и зеленая жабы, чесночница, квакша, жерлянки, прыткая ящерица, несколько видов змей. В Восточной Азии появляются особые виды тех же родов, а также особые роды, например ящерицы-долгохвостки из рода Таchydromus. В Северной Америке вместо семейства ящериц-лацертид (Lacertidae) мы уже находим ящериц-тейид (Teiidae), но зато многие лягушки, жабы и квакши весьма близки к некоторым евразиатским видам. Для Европы характерна огненная саламандра из хвостатых амфибий (Caudata). Несколько других видов этой группы есть также на Кавказе и в Восточной Азии. Однако по богатству хвостатыми земноводными Северная Америка не имеет себе равных.
Большая часть птиц лиственных лесов в летний период преимущественно насекомоядна. Зоофагия свойственна многим из них и зимой. Значительная доля населения представлена перелетными видами - на зиму остается примерно 20-25% от всей численности гнездящихся птиц. Разгар гнездования приходится на май - июнь. В это время численность птиц составляет, как правило, 1-2 тыс. экз. на 1 км2, а биомасса - 30-50 кг/км2. Около 60% населения представлены чистыми зоофагами (насекомоядными и плотоядными видами), около 30% - всеядными, но в летний сезон преимущественно зоофагами и лишь около 10% преимущественно зерноядными птицами. Численность и соотношение разных групп, естественно, могут варьировать даже в одном районе. Так, обычно долинные леса оказываются богаче. Относительно обеднены по общим запасам биомассы и численности наиболее северные варианты неморальных сообществ, особенно те, которым свойственны суровые зимы (например, на Дальнем Востоке нашей страны). Все же главные различия в разных регионах касаются видовых композиций птичьего населения.
Данные по птицам показывают, как средствами совершенно разных фаун в близких экологических условиях строятся во многом сходные сообщества. Такого же рода сравнения для растений оказываются менее показательными, так как между ними наблюдаются, как правило, совпадения на уровне рода. Сравнение хищных млекопитающих во многих случаях выявляет аналоги, относящиеся к разным родам, но состав доминирующих форм у них не столь разнообразен, как у птиц.
Для зверей, представляющих гетеротрофов высших порядков, можно отметить несколько общих черт, которые прослеживаются на всем ареале неморальных сообществ северного полушария. Высокое обилие почвенных беспозвоночных, особенно дождевых червей, позволяет существовать многочисленной группе мелких почвенных и подстилочных зверьков, потребляющих этот корм. Это в первую очередь различные виды семейства кротовых (Talpidae), а также землеройки (семейство Soricidae). В Европе характерен обыкновенный крот Talpa euroраеа, на Дальнем Востоке обычны виды из рода могера (Mogera) и в Японии уротрихус (Urotrichus). В широколиственных лесах Северной Америки кротовые весьма разнообразны, это представители родов Condylura, Scapanus, Scalopus, Neurotrichus. Высокая численность кротов - характерный признак неморальных сообществ. Другие типы экосистем редко имеют аналогичную функциональную группу роющих зоофагов - потребителей крупных личинок и червей почвы.
Общие особенности экосистем
В пределах умеренных широт широколиственные летнезеленые леса (а также и гемигилея умеренно теплых широт) представляют собой наиболее продуктивные сообщества. Особенно это касается чистой первичной продукции, так как сравнительно с тропическими влажными биоценозами потери на дыхание организмов относительно снижаются.
Обычные запасы сухой фитомассы в неморальных ценозах равны 40-50 т/га, или 4-5 кг/м2. Это больше, чем в саваннах и редколесьях, но меньше, чем в гилеях или высоких дождезеленых тропических лесах. Чистая продукция составляет 1-2 ц/га в год, т. е. около 1,5 кг/м2 в год сухой органической массы. Это близко к цифрам для саванн и редколесий средней продуктивности, но уступает продуктивности наиболее богатых высокотравных саванн или редколесных сообществ. Это также близко к данным по продуктивности лавролистных влажных средиземноморских биоценозов (см. карты на с. 62-65). Лавролистные леса и вечнозеленые леса тропических горных стран имеют в своем составе немало гомологов по отношению к неморальной биоте. Без сомнения, ранняя история становления листопадных широколиственных лесов была связана с экосистемами, подобными современным лавролистным сообществам.
Современные листопадные широколиственные леса представляют собой остатки очень широко распространенных уже в самом начале палеогена группировок из листопадных и хвойных пород. Эти палеогеновые леса охватывали сплошной полосой всю северную сушу, занятую сейчас тундрой и тайгой. На месте современных неморальных лесов юга Европы, Кавказа и Передней Азии, Восточной Азии и Восточной Америки располагались почти тропические формации. В течение палеогена и неогена изменения климата приводили к сдвигу на юг природных зон, что сопровождалось перераспределением различных биофилотических элементов. Последнее, в свою очередь, приводило к вытеснению одних видов другими, к формированию региональных различий, которые связаны не только с изменениями климата, но и с различными сочетаниями конкурентных отношений. Поэтому говорить сейчас об «американских», «восточноазиатских» и прочих элементах неморальных ценозов можно лишь с большими оговорками. Значительная часть указанных элементов имеет общий арктотретичный источник, не связанный с каким-либо определенным «центром» возникновения, а развивавшийся на больших пространствах северной суши.
Современные центры разнообразных неморальных флоры и фауны следует рассматривать в первую очередь в качестве центров сохранения, выживания, а не центров формирования.
Много характерных для третичной флоры Северной Евразии и Америки родов сейчас представлено только в Северной Америке и в Восточной Азии (Liriodendron, Nyssa, Aralia, Stewartia, Cephalanthus, Chamaecyparis, Tsuga). Множество третичных видов вымерло на Кавказе и в Европе только к середине плиоцена, хотя часть третичных родов сохранилась в колхидской и гирканской флорах до настоящего времени. Некогда существовавшие в современных неморальных «убежищах» тропические флора и фауна частично сменились «пришельцами», которые не переставали изменяться и на новом месте, частично же преобразовались в умеренно теплые, прибавив убежищам региональную специфику. При этом многие единичные арктотретичные виды, сохранившиеся в убежищах, в прошлом были представлены множеством родственных форм (видов одного рода, секции), входивших в более богатую флору. Это, например, относится к таким родам, как орех, сосна, береза, ольха, бархат, нисса и др.
Богатство видового состава, кроме того, связано и с обширностью территории. Из теории островной биогеографии (см. далее) следует, что ограничение территории автоматически ведет к резкому возрастанию вероятности вымирания. Именно поэтому в сравнительно небольших современных «очагах» неморальных биоценозов не может сохраниться все былое разнообразие, свойственное арктотретичным сообществам. В то же время современная изоляция способствует дивергенции за счет молодых видов, сформировавшихся в течение плейстоцена-голоцена.
Дата добавления: 2019-12-09; просмотров: 1069;
Поиск по сайту
Узнать еще
- F92 Смешанные расстройства поведения и эмоций
- Автомобили-тягачи для вывозки леса
- Анализ процесса качения эластичного колеса по деформируемому грунту
- Антропогенные воздействия на леса и другие растительные сообщества
- Бесстензельная перевозка леса.
- Биологический круговорот масс химических элементов в фитоценозе дубового леса Северной Венгрии
- В продуктах опада деревьев-эдификаторов леса Уокер Бранч,
- Введение в теорию качения колеса

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
