ТУНДРЫ И ПРИПОЛЯРНЫЕ ПУСТОШИ
Географическое распространение,
гигротермический и геохимический режимы
Тундры распространены в Субарктике, образуя циркумполярную зону. В аналогичных широтах южного полушария из-за чрезвычайно ровного морского климата формируются особые сообщества, во многом отличающиеся от тундр. Здесь они названы общим термином «приполярные пустоши». Надо сказать, что в некоторых секторах Субарктики с ровным морским климатом и даже несколько южнее также могут формироваться пустотные сообщества с преобладанием вересковых, осок или некоторых злаков. Следовательно, даже для северного полушария выражение «тундры и приполярные пустоши» оказывается уместным.
Гигротермический режим безлесных приполярных областей показан на приведенных климадиаграммах (рис. 38). Он отличается достаточным или даже избыточным увлажнением, низкими температурами вегетационного периода. Нельзя забывать и об особом режиме освещенности - в середине лета солнце вообще не заходит. В настоящих тундрах период вегетации охватывает короткое летнее время, за которое успевает оттаивать лишь верхний горизонт почв. В субантарктических пустошах вегетационный период может быть растянутым на большую часть года.
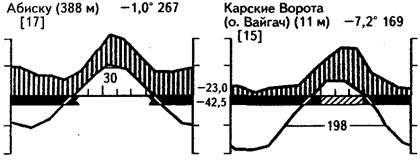
Рис. 38. Климадиаграммы субарктических и арктических районов
Сырые и холодные условия способствуют заболачиванию, а в почвенной толще при этом могут возникать явления оглеения. Весьма распространено также торфообразование, накопление слабо разложившейся органической толщи в условиях избыточного увлажнения и кислой реакции среды. Весьма типичен' мерзлотный микрорельеф (полигональная тундра), а также кочковатость. Профиль глеевых и торфянисто-глеевых тундровых почв из-за мерзлоты неглубокий, дифференцирован он слабо. Даже в верхних горизонтах содержание гумуса не превышает 5 %. Реакция обычно кислая или даже сильнокислая (рН 3,5-5,5). Лишь в южных частях тундры и в лесотундре выражены оподзоленные глеевые и торфянистые почвы.
Сообщества фотоавтотрофов
Севернее границы сомкнутых лесов простирается переходная зона лесотундры, в пределах которой разреженное криволесье, таежные участки по долинам рек и безлесные, поросшие кустарником пространства чередуются друг с другом. Еще севернее простираются так называемые кустарниковые тундры с зарослями ив, карликовых березок (Betula папа), некоторых вересковых. Кустарниковая тундра биогеографически во многом сходна с лесотундрой и северной тайгой. Лишь зона настоящих тундр с мохово-лишайниковым, осоково-пушицевым растительным покровом существенно отличается от таежных массивов.
В зависимости от конкретных местообитаний, в частности от форм мезорельефа, формируются различные сообщества фотоавтотрофов. Каменистые, хорошо дренированные, возвышенные участки имеют разреженный покров из тундрового мятлика (Poa abbreviata),. камнеломки супротиволистной (Saxifraga oppositifolia), дриады точечной (Dryas punctatae). Поверхность камней покрыта накипными лишайниками.
В постоянно увлажненных понижениях сосредоточены гигрофильные группировки с пушицей, осоками, калужницей (Caltha arctica), лютиками (Ranunculus nivalis, R. sulphurous). Наиболее богатые луговидные сообщества приурочены к южным склонам, покрытым слоем мелкозема, здесь господствуют многовидовые сообщества с Cerastium, Draba, Erigeron, Taraxacum. Основные площади плакорных местообитаний покрыты мхами, а в более сухих местах лишайниками. Число видов мхов и лишайников в тундрах достигает тысячи, в то время как цветковых растений насчитывается лишь 200-300 видов. Почвенный покров часто прерывается пятнами голого грунта, морозобойными трещинами и другими формами мерзлотного микрорельефа.
В западной части тундровой зоны Евразии преобладают ягельные тундры с господством видов кладонии (Cladonia), а восточное доминируют виды родов алектория (Alectoria) и цетрария (Cetraria).
Для растительного покрова характерно подавляющее преобладание многолетних видов. Среди них значительна доля очень мелких кустарничков - полярной ивы (Salix arctica), карликовой березки. Очень показательно присутствие вечнозеленых растений: дриады, или куропаточьей травы, вороники, или шикши (Empetrum), брусники. Очень типичны подушковидные жизненные формы, к которым относятся, например, некоторые камнеломки, крупка волосистая (Draba pilosa), подушковидный мак (Papaver pulvinatum). Для областей с морским климатом крайне характерны пустоши с участием разных видов семейства вересковых (вереск, брусника и др.).
Существует мнение, что растения тундры размножаются в основном вегетативным путем. Это полностью справедливо по отношению ко всем растениям, проникающим в тундру из более южных зон (таких видов довольно много). Однако типично тундровые растения периодически, хотя и не каждый год, возобновляются из семян.
В субантарктических регионах в условиях мягкого морского климата существуют своеобразные фитоценозы. При постоянно ветреной туманной и дождливой погоде нередко круглый год днем и ночью температура держится и пределах 0...+5°С. Очень характерны подушковидные растения, например азорелла (Azorella selago) из зонтичных, достигающая в диаметре 1 м. Широко распространены туссоковые злаковники, образующие высокие кочки. Среди них господствуют виды родов Роа и Festuса. Многочисленны разнообразные папоротники (Lomaria alpina, Hymenophyllum peltatum, Polypodium australis, Cystopteris fragilis), плауны (Lycopodium saururus), различные лишайники. Особенно обилен бородатый лишайник Neuropogon melaxanthus. На самом материке Антарктиды обнаружены только два вида цветковых растений - злак Deschampsia antarctica и гвоздичное Colobanthus crassifolius. На острове Кергелен наряду с широко распространенными азореллой и аценой (Acaena adscendens) произрастает крупнолистная «кергеленская капуста» (Pringlea antiscorbutica) из крестоцветных - известное среди моряков противоцинготное средство. На многих субантарктических островах представлены пустоши из вороники красной (Empetrum rubrum) и хебе кипарисовидной (Hebe cupressoides). Местами встречаются участки сфагновых болот. Кроме туссоковых злаковников, характерны злаковые пустоши с преобладанием Cortaderia pilosa. При чрезмерном выпасе злаковники сменяются верещатниками из вороники и представителей родов Blechnum и Pernettia.
В настоящей арктической тундре запасы сухой фитомассы составляют около 5 т/га, причем на многолетние надземные части и корни приходится 4 т/га, а на фотосинтезирующие части - 1 т/га. Годовая первичная продукция составляет 1 т/га. К югу, в направлении к кустарниковой тундре, общая фитомасса достигает 50 т/га, а годовая первичная продукция - 5 т/га. Запасы фитомассы в сообществах Субантарктики также достигают 50 т/га в оптимальных условиях, однако продукция обычно в 2-3 раза выше из-за более растянутого периода вегетации.
Сообщества гетеротрофов
Многочисленные в тундре лишайники по существу являются одновременно и фотоавтотрофами и гетеротрофами. Гетеротрофный компонент этого симбиотического организма, как известно, относится к грибам. Вообще грибы составляют важнейшую часть гетеротрофных уровней в экосистеме тундры (рис.39). Среди микроорганизмов грибы также занимают доминирующее положение по сравнению с бактериями. Недавние исследования микробного населения тундровых почв установили резкие периодические колебания их обилия (раз в 10-15 суток), видимо, не связанные с погодными флуктуациями (вероятно, аутогенные ритмы).
Среди всех животных первое место по суммарной биомассе занимают беспозвоночные, а из них один вид дождевого червя-эйзения Норденшёльда. В моховой дернине в большом количестве встречаются личинки комаров-долгоножек (Tipulidae), а также других двукрылых (Empididae, Muscidae). По численности, как и всюду, преобладают субмикроскопические свободноживущие нематоды (0,5-5 млн экз./м2). Из более крупных червей очень заметны мелкие белые энхитреиды длиной 10-20 мм. Весьма обильны микроартроподы, причем по биомассе обычно преобладают коллемболы из родов Onychiurus, Folsomia, Isotoma. Общая зоомасса почвенных животных в южных вариантах моховых тундр достигает 20 г/м2 (в сырой массе). Примерно половина этого количества приходится на эйзению Норденшёльда, около 2 г - на энхитреид, 1-2 г-на коллембол,
В многочисленных водоемах развивается масса личинок комаров-толкунцов (Chironomidae), а также кровососущих комаров-кулицид (Culicidae).
Упомянутые группы почвенных и водных животных питаются в основном разлагающимися остатками органических веществ. Одними из главных потребителей зеленых растений являются мелкие тундровые грызуны, - лемминги и в меньшей степени полевки. Лемминги и полевки относятся к тем немногочисленным животным, которые деятельны в тундре круглый год. В западных районах Евразии обитает норвежский лемминг (Lemmus lemmus), к востоку от Белого моря его заменяет близкий вид - обский лемминг (Lemmus obensis). Здесь же появляется представитель особого рода - копытный лемминг (Dicrostonyx forquatus). Последний на зиму надевает чисто-белый мех, а два средних когтя передних лап сильно разрастаются, образуя своеобразное «копытце». Этими копытцами лемминг прокладывает подснежные ходы. Только для Америки известен другой вид этого рода - кольчатый лемминг (Dicrostonyx hudsonius). По образу жизни к леммингам близки американские лемминговые мыши (Synaptomys borealis). В берингийской части евразийских и американских тундр среди каменистых россыпей обитают длиннохвостый суслик, или «евражка» (Citellus undulatus), и викарирующие виды сурков (Marmota camtschatica и М. caligata). Северный олень Евразии и американский подвид того же вида, называемый карибу (Rangifer tarandus), обитают в пределах тундровой зоны главным образом в летний период, а на зиму откочевывают в лесотундру. На больших пространствах диких северных оленей заменили стада домашних оленей, но и они ведут образ жизни, сходный с таковым дикого родича. Основной объект питания оленей - ягель (Cladonia, Cetraria) и другие лишайники. Самые крупные стада диких северных оленей живут на Таймыре.
В полярных областях Нового Света до наших дней сохранился реликт ледниковой эпохи - овцебык (Ovibos moschatus). Этих животных успешно акклиматизировали в наших тундрах на Таймыре и на острове Врангеля, а также в тундрах Норвегии и Швеции.
Из птиц наибольшей биомассы на единицу площади достигают различные гуси и казарки, а также белые куропатки. Такое обилие обеспечивается в первую очередь характером питания. Все упомянутые птицы являются потребителями зеленой массы - наиболее доступного и массового корма. Гуси на зиму отлетают далеко на юг, а куропатки откочевывают в лесотундру или скапливаются в заросших кустарниками долинах.
Фоновые виды воробьиных птиц - это пуночка и лапландский подорожник, распространенные циркумполярно. Их следует рассматривать уже как всеядных животных. Преимущественно животные корма потребляют обычные в тундре краснозобые коньки (Anthus cervina) и разнообразные кулики, особенно типичные для тундры песочники (Calidris).

Рис. 39. Основные потоки превратимой энергии в экосистеме тундры
Необходимо хотя бы кратко упомянуть такую специфическую функциональную группу гетеротрофов, как антофилы, которые обеспечивают опыление многих тундровых видов растений. В первую очередь это шмели (Bombus) и мухи-журчалки (Syrphidae). Основу их питания составляют нектар и пыльца. Личинки журчалок живут за счет детрита или хищничают.
Гетеротрофы высших порядков представлены в почвенно-подстилочном ярусе различными многоножками-литобиидами, жужелицами, жуками-стафилинидами, хищными видами нематод и пауками. Из последних особенно заметны пауки-волки из рода Pardosa.
Основные хищники, потребители леммингов, - это песец, белая сова, канюк-зимняк. На водоплавающих птиц и на куропаток охотятся сокол-кречет, ястреб-тетеревятник. Стада северных оленей сопровождает тундровый волк, отбирая в первую очередь молодняк, больных и ослабленных особей. Помимо волка, «потребителями» северного оленя можно считать комплекс кровососов (комары-кулициды и слепни-табаниды), а также разные виды оводов, личинки которых паразитируют в коже (Oedemagena tarandi) и носоглотке (виды рода Oestrus).
Приполярные пустоши Субантарктики из-за островного положения крайне обеднены наземными животными. Особенно это касается позвоночных, которые представлены практически лишь морскими птицами и млекопитающими (трубконосые, пингвины, ластоногие). Характерными птицами субантарктических побережий являются белые ржанки (семейство Chionididae) из отряда куликов. Всего два вида этих птиц (Chionis alba и Ch. minor) держатся на суше, не заходя в воду, и кормятся различными выбросами моря. Внешне они совершенно не похожи на куликов, а скорее напоминают голубей.
Несмотря на изолированность субантарктических островов друг от друга, мелкие почвенные беспозвоночные (нематоды, коллемболы, панцирные клещи и некоторые двукрылые) обнаруживают большую общность видового состава.
Общими чертами приполярных регионов следует считать в основном малое видовое разнообразие, укороченность цепей питания, большие периодические флуктуации численности. Последнее особенно заметно в годы массовых размножений леммингов, с которыми связаны синхронные флуктуации численности пернатых хищников и сдвинутые по фазе колебания обилия песцов. Схемы потоков энергии и структуры трофической сети имеют вид относительно простой системы связей, с сильными облигатными взаимовлияниями (контурами обратной связи). Все это обусловливает слабое развитие компенсаторных механизмов в экосистеме (см. рис. 39).
ВЫСОКОГОРЬЯ
Географическое распространение,
гигротермический и геохимический режимы
Высокогорьем считают субальпийский и альпийский пояса (высотные зоны) горных стран, т. е. территории выше границы сомкнутого леса в тех районах, где выражен горно-лесной пояс. В умеренных и приполярных широтах (начиная примерно от 50°) высокогорья почти идентичны некоторым вариантам расположенной севернее (в северном полушарии) лесотундры и тундры, поэтому далее речь пойдет о высокогорьях умеренно теплых и тропических широт. Эти высокогорья также во многом аналогичны безлесным приполярным территориям, имея нередко общие с ними или близкие виды (арктоальпийское распространение в северном полушарии). Однако здесь уже заметны и различия, которые придают экосистемам низкоширотных высокогорий неповторимые черты. Прежде всего, в отличие от приполярных районов, здесь нет долгого полярного дня и полярной ночи. Фотопериод соответствует широтному положению. Поэтому суточная и сезонная ритмика по мере приближения к экватору становится все своеобразнее. У экватора в высоких горах Южной Америки или в Африке круглый год условия не меняются, каждый день возможны заморозки ночью и сильный нагрев в полдень.
Хотя во всех широтах с подъемом в горы снижаются температуры и обычно хотя бы до некоторого уровня повышается количество осадков (исключая расположенные в дождевой тени склоны), региональные условия горных стран крайне разнообразны и поэтому высокогорья могут быть как засушливыми (Восточный Памир, Тибет, сырты Тянь-Шаня, пуна Анд и т. д.), так и постоянно увлажненными (тропические парамо Анд и высокогорья некоторых районов Африки, Западного Кавказа, Гималаев, Южных Анд). Именно увлажненность в первую очередь приводит к дифференциации условий в горах. При прочих равных условиях засушливость региона приводит к сдвиганию системы высотных поясов вверх. Так, верхняя граница леса на Западном Кавказе и в Альпах нередко проходит на высоте 2000 м абс., а на сухом Восточном Кавказе поднимается до 2700 м абс. В пределах одного региона в Тянь-Шане в зависимости от степени засушливости верхняя граница леса колеблется в пределах 2800-3200 м абс., а нижняя граница языков средних по размерам ледников - в пределах 3000-4000 м абс. Аридные и в особенности семиаридные горные страны в полосе широт 30-45° не имеют сплошных лент высотных поясов, как гумидные горы. Высотные пояса представлены в виде пятен различных по природе экосистем, расположенных на склонах разной экспозиции. Например, в Тянь-Шане на северном склоне хребта Терскей-Алатау, в бассейне реки Чон-Кызыл-Суу, на высоте 2200-2900 м абс., на южных склонах, представлены различные варианты горных степей, а на северных - влажные замшелые ельники из ели Шренка (Picea schrenkiana). В этом районе Тянь-Шаня луговые степи, например, переходя со склонов северной экспозиции внизу на склоны южной экспозиции наверху, встречаются в интервале высот по меньшей мере 1500-3200 м абс.
Сильное влияние экспозиции в семиаридных горных странах субтропических и умеренно теплых широт приводит к тому, что даже при очень резких изменениях климата (например, при понижении температуры в плейстоцене) представленные в горах природные комплексы не исчезают, а лишь перераспределяются по высотам и экспозициям. Такая повышенная буферная способность имеет далеко идущие последствия для состава сообществ, позволяя сохраняться в них древним формам или же закрепляться молодым, недавно возникшим видам. Особенно богаты в этом плане субальпийские группировки. В альпийском поясе условия среды уже начинают приближаться к пределу и состав биоценозов резко обедняется, хотя и включает множество своеобразных по своим адаптациям форм растений и животных.
Невысокие температуры задерживают процессы разложения органического вещества, что приводит обычно к накоплению грубого гумуса и даже к оторфованности верхних почвенных горизонтов. В гумидных условиях это сопровождается сильным выщелачиванием элементов минерального питания и повышением кислотности. При этом нередко возникают пустоши с участием вересковых (рододендроны, верески, вакциниумы и т. д.), осоковых и дерновинных злаков (тассоки Анд, Австралии и Новой Зеландии). Низкие давления, температуры и ветры ведут к физиологическому иссушению даже в достаточно влажных высокогорьях, что способствует широкому распространению подушковидных форм среди растений. Резкие смены погоды, высокий уровень прямой солнечной радиации и ультрафиолетовой радиации приводят к особым биохимическим процессам в живой природе, что отражается и на геохимических особенностях среды. Характерна высокая пространственная дискретность условий вплоть до очень больших различий между куртинами растений и промежутками с редкой растительностью.
Сообшества фотоавтотрофов
Тропические, постоянно влажные высокогорья нередко называют «парамос» - по названию влажных высокогорий в Андах Колумбии, Венесуэлы и Эквадора. Сходные с южноамериканскими по режиму тепла и влаги высокогорья развиты на высоких поднятиях Африки (Камерун, Кения, Килиманджаро, Рувензори, горы Вирунга, некоторые поднятия гор Митумба вдоль рифта), в Индонезии и в Новой Гвинее. Изолированные друг от друга, парамос имеют физиономически сходные растительные сообщества, которые флористически очень различны в тяготеют к ближайшим расположенным ниже участкам. Южноамериканские парамос во многом родственны субантарктическим постоянно влажным безлесным сообществам. Влажные сообщества высокогорий экваториальной Азии имеют в своем составе бореальные элементы и одновременно австралийские. Видимо, такого рода влияния со сдвигом к австралийскому флористическому компоненту имеются в горах Новой Гвинеи, которые пока еще мало исследованы.
Наиболее характерная черта парамос - древовидные растения из преимущественно травянистых родов и семейств. Таковы в Африке гигантские утолщенные крестовники (Senecio johnstonii) и несколько видов лобелий (Lobelia keniensis, L. rhynchopetala, L. deckenii, L. wollastoni и др.) со свечевидными соцветиями. Крестовники относятся к сложноцветным, а лобелии - к колокольчиковым. В парамос Южной Америки растут древовидные эспелеции (Espeletia grandiflora, E. insignia, Е. lopezii) из сложноцветных, называемые «фрайлехонес». Первый из указанных видов обычно имеет высоту около 2 м, но отдельные экземпляры достигают 6 м. Эспелеции внешне похожи на древовидные крестовники Африки. В южноамериканских парамос тоже есть крестовники, но сравнительно низкорослые. Очень похожи своими свечевидными опушенными соцветиями на лобелии гигантские люпины (люпин лисохвостовидный из Эквадора - Lupinus alopecuroides) из бобовых. В экваториальной Азии во влажных высокогорьях также есть древовидные розетковые сложноцветные - анафалисы (Anaphalis). В более сухих горах Южной Америки распространены древовидные розеточные растения - пуйи из бромелиевых. У гигантской пуйи (Puya gigas) из Колумбии агавовидные листья в длину до 1,5 м, короткий стебель, но очень длинный, до 10 м, цветонос. Другие виды имеют толстый стебель и внешне несколько напоминают ксанторею из Австралии. Все древовидные высокогорные растения тропических широт обычно в той или иной степени суккулентны, часто также покрыты густыми волосками и «войлоком» из отмерших листьев.
В парамос обычно хорошо развит моховой (сфагнум), травяной и кустарниковый покров, характерны злаковые кочки, а ниже, у границы с горным лесом, могут расти даже бамбуки. Здесь же развиты кустарниковые и низкорослые древесные разреженные группировки из вересковидных растений (включая как виды семейства вересковых, так и виды других семейств). В Андах характерны рощицы криволесья из видов Polylepis (розоцветные), во всех тропических горах типичны также кустарниковые или древовидные зверобои (Hypericum leucoptychodes в Африке, Н. leschenaultii в Индонезии и Н. laricifolium в Андах) из семейства Guttiferae. Злаковые кочки образованы видами из широко распространенных родов (чаще всего это овсяницы, вейники, бородачи, мятлики, ковыли). Другие травянистые растения включают и красиво цветущие герани, крестовники, лютики, а также различные местные формы, распространение которых ограничено лишь данной горной страной.
Высокогорные подушечники широко представлены не только в тропических, но и в умеренно теплых регионах. Подушки могут быть образованы представителями совершенно разных таксонов, например: из розоцветных - Dryadanthe tetrandra; из свинчатковых - Acantholimon diapensioides; из бобовых -Astragalus в Средней Азии; из зонтичных - Azorella; из кактусовых - Tephrocactus в Андах. Этот список можно было бы легко продолжить за счет видов из самых далеких друг от друга семейств. В сухих нагорьях (пуна Анд, сырты Тянь-Шаня и Памира), кроме подушек, широко представлены дерновинные злаки, которые, однако, не бывают столь высокими, как в парамос. Это опять виды овсяниц, ковылей, мятликов, вейников. В щебнистых участках обычно растут низкорослые суккуленты из толстянковых (очитки Sedum, Rosularia и близкие к ним).
В Южной Америке среди суккулентов большого разнообразия достигают кактусовые, многие из которых очень красиво цветут и высоко ценятся любителями во всем мире (например, виды Lobivia, Pseudolobivia, Rebutia, Oroya, Matucana, Oreocereus, Morawetzia, Mediolobivia, Espostoa и др.). В сухих высокогорьях встречаются также уже упоминавшиеся виды из рода пуйа (включая недревовидные) и другие суккулентные бромелиевые.
В альпийско-гималайском поясе северного полушария в условиях хорошего увлажнения в субальпийском поясе развивается мощное высокотравье, которое перемежается стелющимися кустами и криволесьем из различных древесных растений - это рододендроны, стланики из хвойных (например, арчовый стланик из Juniperus turkestanica), криволесье и низкие кусты из вересковых (кроме уже упомянутых рододендронов). В частности, в горах Азии на юг до Австралии в субальпийском поясе нередко растут кустарниковые и древовидные родичи нашей черники и голубики (например, Vaccinium varingiifolium на Яве). Виды вакциниум обитают также в горах Южной Америки и Мадагаскара. Ареалы многих растений субальпийского пояса позволяют реконструировать пути их расселения и объяснить исторические причины их современной конфигурации.
Сообщества гетеротрофов
Сообщества гетеротрофных организмов, в первую очередь животное население, также представлены разнородными группами, состав которых сильно зависит от регионального положения гор. В условиях сходных гигротермических характеристик среды и у животных можно видеть многочисленные примеры параллелизмов и конвергенции наряду с уникальными чертами облика и экологии в каждом районе. Таксономические связи с удаленными территориями чаще полнее проявляются у беспозвоночных животных, особенно у самых мелких их представителей - нематод и микроартропод. Наиболее своеобразен состав населения позвоночных, в первую очередь зверей и птиц.
Крупные травоядные млекопитающие, характерные для альпийско-гималайской полосы горных стран, - это представители нескольких видов козлов (Capra) и баранов (Ovis). В Кордильеры Северной Америки проникает снежный баран, живущий и у нас в горах Сибири, но одновременно там появляется чрезвычайно своеобразная снежная коза (Oreamnos americanus) - белого цвета, с маленькими рогами и гривкой на холке. В Евразии в отдельных горных странах распространены и другие копытные - серны, тары, горалы, серау, голубые бараны, яки. В парамос Южной Америки обитают редкие шерстистые, или горные тапиры (Tapirus pinchaque) и мелкие высокогорные олени (Pudu mephistopheles). В парамос и в более сухих высокогорьях (пуна, халка) встречаются и другие виды оленей, из которых перуанский олень (Hippocamelus antisiensis) держится среди скал и напоминает повадками горного козла. В пуне характерны и безгорбые верблюды из рода лама. Все горные травоядные часто заходят в верхний пояс горного леса, для них характерны периодические вертикальные миграции. Это же можно сказать и о хищных млекопитающих - различных медведях, лисицах, кошачьих. Напротив, мелкие грызуны ведут строго оседлый образ жизни, но у них вырабатываются особые приспособления против сезонных и даже суточных колебаний непостоянной погоды высокогорья - устраивают хорошо защищенные убежища в норах и среди камней, запасают сено, впадают в спячку в холодный период.
В умеренно теплых широтах у птиц нередко возникает весьма узкая кормовая специализация, невозможная в равнинных условиях. Например, ряд видов горных зерноядных птиц Средней и Центральной Азии (Leucosticte, Serinus pusillus) в течение нескольких месяцев могут поедать семена немногих видов растений, ежедневно опускаясь из высокогорий в теплые долины ранней весной. В течение лета эти растения созревают на все больших высотах, постепенно доходя до высокогорий. Ближе к осени в долинах созревает второй урожай, позволяя горным вьюркам не менять кормового рациона. Таким же образом ведут себя и горные виды колибри в умеренных широтах, передвигаясь вместе с движением в горы волны цветения. Напротив, в ровных условиях влажных экваториальных высокогорий те же колибри и заменяющие их в Старом Свете нектарницы передвигаются не столько в вертикальном, сколько в горизонтальном направлении, отыскивая цветущие растения, которые появляются в любое время года.
Другие виды птиц в течение неблагоприятного сезона мигрируют в долины и там проводят некоторое время, часто резко изменяя характер корма. Например, обитающая в летнее время поблизости от ледников и поедающая насекомых краснобрюхая горихвостка (Phoenicurus erythrogaster) зимует в долинах с зарослями облепихи и питается в основном ягодами этих кустарников (краснобрюхая горихвостка живет на Кавказе, в Средней и Центральной Азии). Весьма характерны во многих горах и главным образом растительноядные формы, поедающие в большом количестве зеленую массу альпийских трав, которая очень богата протеином и усвояемыми углеводами и жирами. Таковы, в частности, улары (poдTetraogallus). Наконец, в составе населения всегда представлены всеядные птицы (например, альпийские галки Евразии) и крупные хищники. Среди последних многочисленны падалыцики - грифы, сипы, бородачи в Старом Свете, кондоры в Новом Свете. Крупные хищники подолгу парят в небе, используя восходящие потоки воздуха, всегда имеющиеся в горах.
Состав даже наиболее массовых птиц очень различен в разных регионах, ведь некоторые горные виды обладают крошечными ареалами, как, например, некоторые колибри Южной Америки. Все же можно для высокогорий Старого Света, кроме уже упомянутых альпийских галок (Pyrrhocorax graculus), грифов и бородачей, горных вьюрков (Leucosticte), отметить еще горных коньков (Anthus spinoletta), рогатых жаворонков (Eremophila alpestris), снежных вьюрков, или снежных воробьев (Montifringilla), ряд видов горных чечевиц (Carpodacus), стенолаза (Tichodroma muraria). Из них горные коньки и рогатые жаворонки широко распространены в горах Северной Америки, при этом рогатые жаворонки населяют также тундру и пустынные низкогорья Центральной Азии. В изолированных горах Африки для пояса древовидных сенецио и лобелии характерна нектарница Джонсона (Nectarinia johnsoni).
В парамос Анд обильны виды, многие из которых встречаются у уровня моря на южной, холодной оконечности материка. Среди фоновых видов это, например, водяной печник (Cinclodes fuscus), пищуховый землелаз (Upucerthia) из семейства печников (Furnariidae) и скальный земляной тиранн (Muscisaxicola alpina) из семейства тиранновых. Обычны также дрозды Turdus fuscater, два вида крапивников и несколько видов овсянковых, танагр и колибри. Из последних наиболее обычны крошечные Metallura и Oxypogon, которые поднимаются в горы на 5000 м абс. и часто кормятся среди листьев и цветов древовидных эспелеций (фрайлехонес). В пуне к фоновым видам принадлежит широко распространенный в Новом Свете вид овсянковых - зонотрихия (Zonotrichia capensis), некоторые тинаму, а также похожий на поползня кроличий землекоп, или «шахтер» Geositta cunicularia из семейства печников. Как и в сухих высокогорьях Центральной Азии, в пуне многие птицы гнездятся в норах и нишах обрывов. Это упомянутый «шахтер» и многие другие виды печников. Многие из них сооружают гнезда из глины, за что и получили такое название. Андский шилоклювый дятел (Colaptes rupicola) образует колонии и сооружает норы в обрывах, наподобие наших ласточек-береговушек. Ближайший родич этого дятла живет в лесах Северной Америки. В пуне обычны также некоторые виды колибри и мелкие горлицы.
Дата добавления: 2019-12-09; просмотров: 1345;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
