И КОЛЮЧИЕ КСЕРОФИТНЫЕ ЗАРОСЛИ
Географическое распространение,
гигротермический и геохимический режимы
В настоящем разделе рассматриваются весьма разнообразные сообщества, объединяемые в первую очередь по общему признаку засушливости, выраженной большую часть года. Понятие засушливости, или аридности, предполагает в самой общей форме дефицит влаги в экосистеме. Степень аридности оценивается с помощью различных коэффициентов, учитывающих соотношение прихода и расхода влаги. Для этой цели в разное время были предложены различные формулы расчета. Так, например, широко известен коэффициент увлажнения В.В. Докучаева - Г.Н. Высоцкого - Н.Н. Иванова:
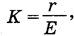
где в числителе - годовая сумма осадков, а в знаменателе - потенциальная годовая испаряемость с открытой водной поверхности. По мере увеличения аридности коэффициент К уменьшается.
В последнее время большинство географов и экологов предпочитает использовать радиационный индекс сухости М.И. Будыко:
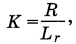
где R - годовой радиационный баланс, L - скрытая теплота испарения, г- годовая сумма осадков. Величина индекса, близкая к единице, характеризует гигротермические условия, наиболее благоприятные для развития высокопродуктивных биомов. В умеренных широтах - это широколиственные леса и лесостепи, в субтропических - гемигилеи, в тропических и экваториальных - низинные дождевые леса (истинные гилей). К югу от лесостепи индекс увеличивается. Индекс в пределах от 1,2 до 3 определяет развитие степных или жестколистных средиземноморских сообществ. Индекс от 3 до 5 соответствует гигротермическим условиям полупустынь умеренных широт (пустынных степей), полупустынь Патагонии и крайних по засушливости вариантов средиземноморских сообществ (фриганы, сухие варианты первичного шибляка, как фисташковое редколесье или миндальники, предгорные эфемеровые сообщества Средней Азии и Ближнего Востока, заросли держидерева и т. п.) и некоторых вариантов тропических саванн. И наконец, индекс более 5 характеризует пустыни как умеренных, так и тропических широт и различные типы тропических колючих ксерофитных зарослей (буш Сомали, каатинга Бразилии, малга-скраб Центральной Австралии, южные районы Гран-Чако). В наиболее засушливых (экстрааридных, или гипераридных) областях земного шара величина указанного индекса может достигать 50 и более. -
Известная карта ЮНЕСКО «Аридные области земного шара» первоначально составлена П. Мейгсом в 1952 г. на основе индекса влажности, предложенного климатологом Торнтвейтом:
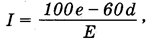
где е - избыточная влага осадков во влажные месяцы (превышение над месячной суммой 100 мм), d - дефицит влаги в сухой период, Е - потенциальная эвапотранспирация. Для семиаридных областей индекс Торнтвейта колеблется в пределах от -20 до -40, а для аридных всегда оказывается меньше -40.
Различные подходы к оценке степени засушливости определяются тем, что нелегко выделить универсальный критерий и абсолютно ведущие факторы, от которых зависит явление аридности. Однако во всех случаях пользоваться одной лишь нормой годовых осадков явно недостаточно, хотя даже и такой чрезвычайно упрощенный подход дает в первом приближении удовлетворительные результаты (например, нередко определяют границу пустынь и полупустынь в пределах умеренных широт по изогиете 200 мм).
Для полной и дифференцированной характеристики различных типов аридных экосистем необходимо анализировать сезонную ритмику температур и увлажнения с учетом соотношения и крайних значений этих параметров. Разнообразие гигротермических режимов различных аридных областей земного шара иллюстрируют приведенные климадиаграммы (рис. 25).
Первый, наиболее выраженный тип аридных экосистем формируется в условиях полного или почти полного отсутствия осадков. Такие экосистемы мы находим, например, в Центральной и Восточной Сахаре (см. рис.25, а, б - Адрар и Асуан). В пустынях этого типа плакорные участки практически лишены жизни, которая сосредоточена лишь в глубоко врезанных долинах и у выходов грунтовых вод. Пожалуй, это единственный тип пустынь, полностью оправдывающих свое название. Впрочем, и здесь несколько раз в столетие случаются дожди, после которых, казалось бы, безжизненная равнина покрывается скудной зеленью из проросших семян.
Пустынные экосистемы второго типа также практически лишены дождей, однако некоторое количество влаги поступает в экосистему в виде конденсата частых туманов. Речь идет о прибрежных тропических и субтропических пустынях Атакама (см. рис. 25, в) и Намиб (см. рис. 25, г). В отличие от пустынь первого типа прибрежные пустыни туманов имеют значительно более ровный годовой ход температур на более низком общем уровне. В результате среднегодовые температуры обычно не превышают +20°С. Увлажняющее воздействие тумана заметно прежде всего на наветренных склонах и гребнях. Именно здесь можно видеть разреженный растительный покров, включающий специфические жизненные формы растений - конденсаторов влаги туманов, а в понижениях и на выровненных участках растительность отсутствует.
В тропических и субтропических областях с летним максимумом осадков формируется третий тип аридных экосистем. Примером может служить малга-скраб Центральной Австралии (см. рис. 25, д) или Южная Сахара (см. рис. 25, е). К этому же типу примыкает полоса Сахели, по характеру гигротермического режима образующая переходную зону к типичным саваннам и редколесьям.
Выровненный ход условий вегетации благоприятствует развитию разреженных редколесий из различных видов акаций. Примечательно, что виды этого рода распространены на всех материках и входят в состав эдификаторов в большинстве аридных сообществ данного типа.
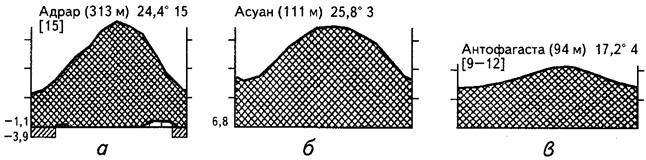
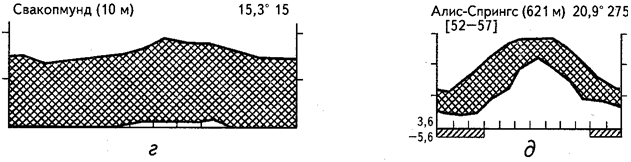
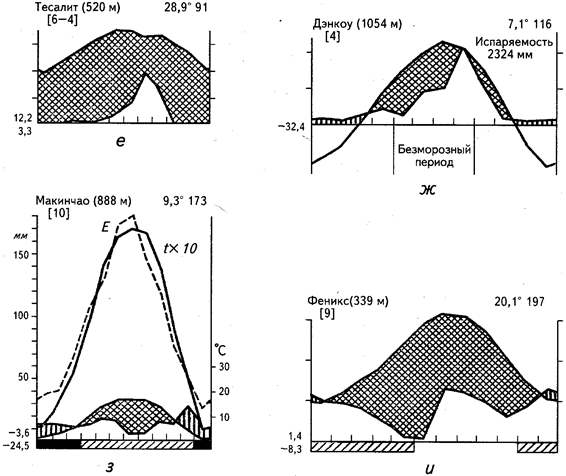
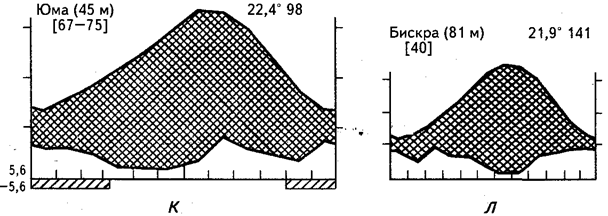
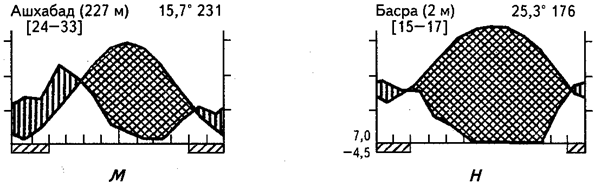
Рис. 25. Климадиаграммы аридных районов
Летний максимум осадков наблюдается также в аридных экосистемах четвертого типа, однако они хорошо отличаются наличием холодного зимнего периода. Отсюда ясно, что экосистемы данного типа приурочены к умеренным широтам. Примером могут служить пустыни центральноазиатского типа, представленные в Казахстане и Центральной Азии (см. рис. 25, ж). Холодный (а в ряде районов и морозный) период обусловливает задержку вегетации. В растительном покрове преобладают длительно вегетирующие ксерофитные полукустарники (полыни, солянки, терескен и т. п.) и кустарники (карагана, эфедра). Характерно почти полное отсутствие эфемеров.
Пятый тип аридных экосистем представляет полупустыня Патагонии, имеющая весьма сходную с предыдущим типом структуру растительного покрова (см. рис. 25, з). Весьма значительны различия в климадиаграммах. Однако, несмотря на это, ритмика вегетации аналогична, что определяется в первую очередь наличием морозной зимы. Именно по этой причине растительность вегетирует летом, несмотря на сравнительно жесткие условия увлажнения.
В тропических и субтропических широтах формируется шестой тип аридных экосистем, включающий весьма разнообразные варианты. Объединяющим признаком может служить нерегулярный или двувершинный ритм увлажнения в сочетании с благоприятным температурным режимом, дающим возможность зимней вегетации.
К этому типу относятся Сонорская пустыня (см. рис.25, и, к), некоторые районы Сахары (см. рис. 25, л), полупустыни Верхнее и Большое Карру, колючие ксерофитные заросли Сомали, Гран-Чако, бригелоу-скраб Северо-Восточной Австралии. В растительном покрове экосистем этого типа весьма характерны древесно-кустарниковые формации, различные суккуленты, а также геофиты.
В последний, седьмой тип аридных экосистем входят субтропические и умеренно теплые пустыни, низкотравные полусаванны, суккулентные и колючекустарниковые заросли, существующие в условиях влажного зимнего периода и длительной летней засухи. В пределах Евразии это ирано-туранские пустыни и низкотравные полусаванны: эфемеровые травянистые сообщества предгорий (см. рис. 25, м, н). Сходные условия наблюдаются и в других регионах земного шара при переходе от экстрааридных пустынь (экосистемы первого-второго типов) к жестколистным формациям средиземноморского климата. На рис. 21 можно наглядно проследить экотон от пустыни второго типа к жестколистному средиземноморскому маторралю. В переходной зоне формируется пустыня седьмого типа. Аридные суккулентные сообщества с выраженным периодом зимних дождей представлены также в Малом Карру (Капская провинция). Наиболее характерным элементом сообществ в аридных экосистемах седьмого типа являются эфемеры и эфемероиды. В тех вариантах, которые формируются в условиях мягкого зимнего периода, возможно широкое развитие суккулентов, подобно шестому типу, на песках обычны пустынные кустарники и редколесья. Такие варианты образуют постепенный переход между экосистемами шестого и седьмого типов.
Геохимические особенности аридных областей, несмотря на разнообразие гигротермических условий, формируются под влиянием одного ведущего фактора - полного или почти полного отсутствия промывного режима. Это определяет широкое распространение засоленных почв. Количество гумуса колеблется весьма сильно, прямо коррелируя с продуктивностью биоценозов. Однако при всех возможных колебаниях содержание гумуса даже в самых верхних горизонтах редко превышает 2-3%. В то же время запас питательных веществ в почвах может быть достаточно высоким, и при орошении это позволяет собирать большие урожаи. Однако в этом случае резко возрастает опасность накопления воднорастворимых солей (в первую очередь хлоридов натрия и магния). Реакция среды в почвах аридных территорий обычно сдвинута в щелочную сторону или близка к нейтральной. Кроме воднорастворимых солей, относительно накапливаются соединения карбонатов кальция и магния, а также слаборастворимого гипса (сернокислого кальция).
Эдафические факторы играют большую роль в классификации и типологии пустынных экосистем. Свойства субстрата оказывают подчас непосредственное влияние на распределение и численность как растительных, так и животных организмов. Поэтому вполне закономерно, что именно пустынные экосистемы часто типизируют и называют соответственно характеру субстрата. Так, выделяют пустыни песчаные, глинистые, лёссовые, каменистые (галечниковые и щебнистые), скальные (рис. 26). По более общим геоморфологическим характеристикам различают два основных типа пустынь - аккумулятивные и денудационные. В первых накапливаются большие толщи рыхлых аллювиальных и пролювиальных отложений. Песчаные отложения в аккумулятивных пустынях могут переотлагаться ветром, образуя эоловые формы, такие, как грядовые, бугристые, ячеистые и барханные пески. Денудационные пустыни относительно подняты - это плато и нагорья с обнажением коренных пород, а иногда и с развитой корой выветривания (элювиальный плащ). По склонам возвышенностей формируются делювиально-пролювиальные отложения. Среди аккумулятивных пустынь по характеру субстрата преобладают песчаные и глинистые, а в типе денудационных - щебнистые и скальные.
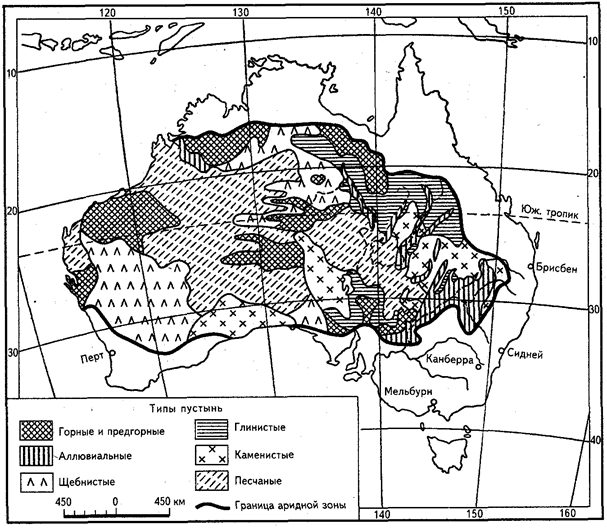
Рис. 26. Типология пустынь Австралии по эдафическим и геоморфологическим характеристикам (по Р. Слэтьеру и Р. Перри, 1969)
Сообщества фотоавтотрофов
Общий облик сообщества фотоавтотрофов аридных областей характеризуется в первую очередь разреженностью, несомкнутостью надземных частей фитомассы. В то же время на единицу площади транспирирующей поверхности как в гумидных, так и в аридных областях приходится почти одинаковое количество осадков. Таким образом, расход воды на единицу площади листовой поверхности остается примерно одинаковым. Во всех засушливых областях с увеличением аридности относительно увеличивается доля подземной фитомассы. Исключением являются лишь сообщества с преобладанием суккулентов. Одновременно с увеличением аридности растительный покров все более сосредоточивается в отрицательных формах рельефа - руслах временных водотоков (вади), в эрозионных ложбинах и котловинах. Рыхлые субстраты, как правило, лучше обеспечивают влагой зеленые растения. Поэтому при прочих равных условиях растительный покров на песчаных участках всегда значительно богаче и разнообразнее, чем на тяжелых глинистых субстратах.
В пустынных экосистемах первого типа часто на огромных пространствах вообще невозможно обнаружить и единичных экземпляров высших растений. И даже в тех местах, где по руслам вади произрастают отдельные особи, они не формируют растительного покрова. Видовой состав крайне беден. Так, например, в Центральной Сахаре в окрестностях оазиса Феззан встречаются лишь парнолистники Zygophyllum simplex, Z. album, Fagonia bruguieri, Cornulaca monacantha, Traganum nudatum, Aristida pungens и верблюжья колючка Alhagi camelorum.
Пустыни второго типа ненамного богаче, однако в отдельных участках там все же формируется растительный покров, хотя и очень разреженный. В прибрежной части пустыни Намиб преобладают песчаные и в особенности щебнистые грунты. Увлажняемые туманом участки покрыты пестрыми кустистыми, листоватыми и накипными лишайниками. В годы с дождями развиваются однолетние эфемерные суккуленты Hydrodea bossiana. Местами встречаются также плотные подушки уже упомянутого парнолистника (Z. simplex). Кроме того, на этих участках произрастают единичные особи суккулентов из семейства Aizoaceae. В сухих засоленных долинах можно видеть небольшие кустики безлистного многолетника Arthraerua из семейства амарантовых. На расстоянии более 50 км от берега характер растительности меняется, появляются сообщества с доминированием злаков из рода аристида (Aristida), здесь наблюдается переход к пустыне третьего типа.
В пределах полосы туманов наиболее разнообразны сообщества суккулентов на скалах и щебнистых наносах. Это стеблевые (Trichocaulon, Hoodia, Senecio longiflorus, Euphorbia brachiata) и листовые (Lithops и Hereroa) суккуленты (вторые из семейства Aizoaceae) и Aloe asperifolia из семейства лилейных. Наиболее многочисленны на скалах лишайники. Упомянутые литопсы представляют собой типичный пример «оконных» растений.
Большая часть этого миниатюрного, длиной всего 3-4 см, растения погружена в субстрат. На поверхность выходит лишь плоская, как бы срезанная верхушка утолщенного листа растения, покрытая прозрачной кутикулой. Сквозь это своеобразное «окно» свет попадает к фотосинтезирующим органам литопса. Таким образом достигается крайняя степень экономии влаги путем снижения транспирации (рис. 27).
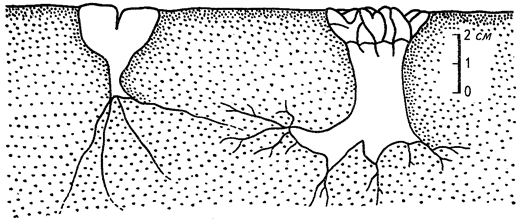
Рис. 27. Корневые системы литопса (Lithops, слева) и налантуеа (Nananthus, справа) (по Г. Вальтеру, 1968)
Рис, 28. Корневая система питурантуса скрученного (Pituranthus tortuosus)
(по Каушу, из Г. Вальтера, 1968)
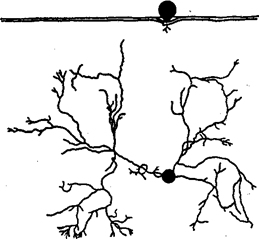
Рис. 29. Корневая система ферокактуса (по Г. Вальтеру, 1968):
вверху — профиль, глубина проникновения корней — 2см; внизу — план
Разнообразие корневых систем растений пустынь крайне велико (рис. 28) и связано вариантами перехвата атмосферной влаги.
Наиболее известное растение в пустыне Намиб - вельвичия удивительная (Welwitschia mirabilis) из семейства Welwitschiaceae голосеменных, растущая отдельными экземплярами в верховьях русел сухих водотоков. В ее корневой системе удачно сочетаются длинные корни, проникающие в глубину до 1,5 м, использующие влагу редких дождей, и боковые корни, ветвящиеся у самой поверхности и улавливающие влагу туманов. Поверхностный характер строения корневой системы имеют ферокактусы, обитающие в пустыне Сонора (рис. 29).
Аналогом пустыни Намиб на Тихоокеанском побережье Южной Америки является пустыня Атакама. В прибрежной полосе растительность образована отдельными куртинами бромелиевых из рода тилландсия (Tillandsia). Эти растения поглощают росу не корнями, а листьями, с помощью чешуевидных волосков. Местами поверхность на значительных площадях лишена высших растений, и лишь сине-зеленые водоросли и кое-где кустистые лишайники рода Cladonia (ягели) покрывают бесплодную почву. Выше по склонам гор увлажняющее влияние туманов увеличивается, здесь появляются различные маревые, гвоздичные, пасленовые, злаки и в особенности суккулентные представители кактусовых и молочайных. Украшением пустынных равнин Атакамы на севере Чили являются куртины кактусов Copiapoa cinerea с черными, как смоль, шипами и белой, словно покрытой мелом, верхушкой.
Тропические пустыни и полупустыни третьего типа представляют собой акациевые редколесья в сочетании с крайне разреженными злаковниками и пустынными кустарниками. В Центральной Австралии такого рода сообщества именуют малга-скраб. Малга - это акация Acacia aneura, у которой функции листьев выполняют филлодии - расширенные черешки. Кроме этого доминирующего вида, в состав пустынных редколесий входит еще несколько видов филлодийных акаций (рис.30), а также казуарина (Casuarina lepidophloia и другие виды). Вдоль сухих русел встречаются толстые невысокие эвкалипты (Eucalyptus camaldulensis), а в более благоприятных условиях (обеспеченные грунтовой водой русла) - высокие с красивыми белыми стволами эвкалипты-привидения (Е. рариапа). В этих галерейных зарослях участвуют также Е. dichromophloia. Acacia estrophiolata, Melaleuca ssp. На плакорных песчаных участках плотные подушки образуют злаки Triodia pungens и Т. basedowi, несколько реже - виды рода Plectrachne. В массивах грядовых песков куртины злаков занимают межгрядовые понижения. Там же, в понижениях, обычны отдельные виды разнотравья и кустарники: протейные (Hakea leucoptera, Grevillea stenobotrya и G. juncifolia), представители семейства Myoporaceae (Eremophila longifolia и E. latrobei), маревые (Rhagodia spinescens и Euchylaena tomentosa), молочайные (Euphorbia wheeleri, Adriana hookeri).
Рис. 30. Различные австралийские акации с филлодиями
Выше по склонам песчаных гряд произрастают кроталярии (Crotalaria cunninghami, С. dissitiflora), гребневую часть занимают единичные Zygochloa paradoxa и многолетники Ptilotus latifolius из амарантовых и Sida corrugata из мальвовых. После редких дождей поверхность песка покрывается нежной зеленью эфемеров (Salsola kali, Plagiosetum refractum, Trichodesma zeylanicum).
В местах с засолением, особенно по краям высохших озер, типичны Nitraria schoberi (этот же вид встречается в пустынях Евразии), виды маревых из родов лебеда (Atriplex) и марь (Chenopodium). На плотных глинистых грунтах с засолением образуется покров из Atriplex vesicaria и Kochia sedifolia. Такие же растительные группировки характерны и для расположенной южнее полупустыни Налларбор.
В пустынных районах южной периферии Сахары, где первый тип аридных экосистем сменяется третьим, скудные осадки выпадают летом. Редко разбросанные куртины злаков Aristida и Andropogon перемежаются с обширными участками обнаженной почвы.
При количестве осадков в пределах 50-100 мм в год по дну сухих русел (вади) появляются акациевые редколесья из Acacia flava, A. orfota, A. tortilis. В районе Хартума на песчаных почвах (150 мм осадков в год) Acacia tortilis выходит на плакорные участки, где образует крайне разреженные сообщества. На таких почвах при осадках в 200-300 мм (зона Сахели) произрастает также A. mellifera, появляются первые экземпляры A. Senegal и A. seyal, характерных в основном для типичных саванновых редколесий.
В аридных участках третьего типа в пределах Южной Африки (полупустыня Калахари) одним из самых распространенных злаков является аристида одноперистая (Aristida uniplumis). Аридные редколесья представлены несколькими видами акаций, в частности Acacia detinens. В более засушливых участках древесная растительность практически исчезает, злаковый покров становится все более разреженным, появляются отдельные суккуленты (молочаи, алоэ, различные толстянковые), кустарники, среди которых обычны парнолистники (Zygophyllum) и дереза (Lycium). Так при дальнейшем движении на юг можно наблюдать постепенный переход к аридным экосистемам шестого типа, представленным полупустынями Верхнее и Большое Карру.
Аридные экосистемы четвертого типа представлены в самых северных вариантах пустынь и полупустынь Евразии и Северной Америки. Растительный покров сложен преимущественно многолетними полукустарниками: полынями (Artemisia), маревыми (Salsola, Anabasis, Kochia, Reaoumuria, Sympegma). Весьма характерны также невысокие кустарники (Caragana, Nitraria, Calligonum). В некоторых местах доминирующее положение в растительном покрове занимают кусты эфедры (Ephedra), терескена (Eurotia), саксаула (Haloxylon). Злаки обычно представлены мелкодерновинными формами, например галечниковым ковылком (Stipa gloriosa). В полупустынях Евразии, как и в сухих степях, обычен также типчак (Festuca sulcata). Следует иметь в виду, что в Северной Америке наиболее сухие варианты короткотравной прерии по существу представляют собой полупустыню с господством полыней, мелкодерновинных злаков и небольших стелющихся опунций (Opuntia). В пустынях Большого Бассейна характерны сообщества из солянок и крупных кустов полыни трехзубчатой (Artemisia tridentata). Местами обычен также североамериканский вид терескена (Eurotia lanata).
В англоязычной литературе варианты аридных экосистем, таких, как полынные и солянковые полупустыни, именуются степями (dry steppe, bush-steppe). Это следует учитывать при работе с иностранными источниками. В русском языке, хотя и не в научной литературе, также нередко используют слово «степь» в названиях отдельных полупустынных и пустынных регионов (Муганская степь, Мильская степь. Голодная степь, Чуйская степь).
Аридные экосистемы пятого типа, представленные полупустынями Патагонии, также часто неправильно именуют степями. В растительном покрове Патагонии доминируют кустарнички, часто образующие подушковидные формы. Особенно характерны большие колючие подушки зонтичного Melinum spinosum. В более увлажненных западных районах Патагонии обычны низкотассоковые (мелкокочковатые) злаки Festuca pallescens и Stipa speciosa. Кстати, оба эти вида произрастают также в Новой Зеландии. В центральной части региона большие площади занимают сообщества с господством сложноцветных (Nassauvia glomerulosa, виды родов Chuquiraga, Grindelia, Senecio). Характерно также присутствие эфедры (Ephedra) и Adesmia campestris из бобовых. В более влажных и прохладных южных районах появляются субантарктические азореллы и ацены (Azorella, Acaena). На каменистых солнечных участках в изобилии встречаются куртины мелких кактусов (Maihuenia patagonica, виды родов Austrocactus, Tephrocactus, Opuntia). В наиболее теплых северо-западных районах Патагонии появляются крупные, высотой до 2 м, кусты Prosopis denudans и Larrea nitida. У западной границы, на переходе к лесам из южного бука, характерны протейные (Embothrium coccineum с яркими красными цветами и другие виды этого семейства), а также различные виды барбариса, смородины.
Растительный покров аридных экосистем шестого типа с благоприятным режимом температур (отсутствие морозного периода) и нерегулярными осадками обычно представлен зарослями колючих кустарников и суккулентов. Нередко хорошо выражены также аридные редколесья. Злаковый покров развит слабо. В Новом Свете сообщества этого типа представлены на северо-западе Аргентины, в Мексике и на юге США. Как в Южной, так и в Северной Америке весьма типичны сообщества с доминированием видов рода Larrea из парнолистниковых. В Аргентине и Парагвае произрастает L. nitida, а в Северной Америке - L. tridentata (креозотовый куст). В обоих указанных регионах представлены и другие викарирующие виды из одних и тех же родов (Prosopis, Opuntia, Ephedra, Acacia, Atriplex). В обоих районах сосредоточено также множество (более 2 тыс. видов) кактусовых. В северном очаге произрастают гигантские, высотой до 15 м, «сагуаро» (Carnegia gigantea), мексиканские «вьехос» (Cephalocereus senilis), достигающие почти такого же размера, как и сагуаро; бочковидные, вооруженные громадными шипами виды родов Ferocactus, Echinocactus, мелкие виды родов Astrophytum, Mamillaria, Echinocereus, Lophophora, Coryphanta. Для южного очага характерны столбовидные, высотой до 10 м, виды рода Cereus, мощные двухметровые Helianthocereus, множество мелких видов Parodia, Gymnocalycium, Rebutia, Lobivia. Особенно велико разнообразие кактусов в предгорных и горных аридных экосистемах.
Между полупустыней Калахари (третий тип) с летним периодом дождей и Малым Карру с зимним периодом дождей (седьмой тип) расположены аридные области: Верхнее Карру с нерегулярными осадками и Большое Карру с осенними осадками. Обе эти области, особенно Верхнее Карру, относятся, как и пустыня Сонора, к шестому типу. Для них особенно характерно большое участие в растительном покрове суккулентов. В верхнем Карру наиболее типичны сообщества с доминированием кустарничков из семейства сложноцветных (Pentzia, Pteronia, Eriocephalus, Chrysocoma, Euryops, Diplopappus). Участием сложноцветных кустарничков сообщества Верхнего Карру в некоторой степени сходны с полупустынями Патагонии (см. выше). Этому сходству способствуют также частые зимние похолодания, вследствие значительной высоты над уровнем моря (1100-1300 м). Однако весьма характерно участие листовых суккулентов из семейства аизооновых (Aizoaceae) Mesembryanthemum, Lithops. Характерны также луковичные и клубневые геофиты из лилейных, амариллисовых и ирисовых.
Особенно богата суккулентами область Большого Карру. Кроме листовых суккулентов из аизооновых (Fenestraria, Lithops, Pleiospilus, Faucaria), представлены также виды толстянковых (Crassula и др.), сложноцветных (Senecio), лилейных (Aloe, Haworthia, Gasteria). Среди стеблевых суккулентов преобладают молочайные (многочисленные виды Euphorbia) и ластовниевые (Asclepiadaceae) - Stapelia, Huernia, Hoodia, Trichocaulon, Caralluma). На засоленных участках сосредоточены заросли тамариска, солянок (Salsola), лебеды (Atriplex halimus).
Длительная летняя засуха и зимние осадки характерны для аридных экосистем последнего, седьмого, типа. К этому типу, в частности, относится лежащее южнее Большого Карру аридное нагорье Малое Карру. Здесь представлены почти все группы суккулентов, перечисленные для Большого Карру. Еще большего развития достигают геофиты-эфемероиды из лилейных, амариллисовых, ирисовых и кисличных. Весьма заметна также примесь типично капской склерофилльной флоры, включающая виды протейных, масличных. В растительном покрове господствуют невысокие кустарники в суккуленты (Carissa ferox, Dodonaea thunbergii, Euclea undulata. Cotyledon, Elytropappus, Asparagus, виды Aizoaceae). Северная субтропическая часть Сахары с зимними осадками также относится к седьмому типу аридных экосистем. Наиболее богата растительность песчаных пустынь (эргов). Как и в Центральной Сахаре, здесь обычна верблюжья колючка, часто образует разреженный злаковый покров Aristida pungens. Очень разреженный древесно-кустарниковый ярус сложен сахарским дроком (Genista saharae), эфедрой крылатой (Ephedra alata), тамариском (Tamarix articulata), Calligonum comosum, Puturanthos tortuosus, Retama retam. В межбарханных понижениях растут сыть пустынная (Cyperus conglomeratus), злак дантония ломкая (Dantonia fragilis).
Вдоль русел вади, кроме упомянутых растений, можно обнаружить Zilla spinosa, Zygophyllum, Zizyphus, Balanites, Pistacia, Acacia. Древесные формы поселяются здесь, используя временный сток. Около выхода грунтовых вод развиваются оазисы из пальмы дум (Hyphaene thebaica), акаций, каперсов. В окультуренных оазисах эта естественная растительность вытеснена культурой финиковой пальмы (Phoenix dactylifera).
Щебнистые пустыни обычно имеют очень разреженный покров из подушковидных и колючих растений, например Fredolia aretioides из маревых, Limoniastrum feei из свинчатковых. Весьма примечательна иерихонская роза (Anastatica hierohuntica) из крестоцветных. Во время засухи побеги с созревшими плодами загибаются вверх и внутрь, образуя плотный шар. Нередко он отрывается от корня и переносится ветром. С первым дождем набухающие ветви распрямляются как пружина и разбрасывают вокруг семена.
На восточной периферии Сахары к юго-востоку от Каира выделяется несколько типов сообществ. На плакорах среди песчано-каменистых участков доминируют кусты саксаула (Haloxylon salicornicium). В пролювиальных ложбинах, кроме того, в большом количестве представлены куртины Panicum turgidum и Lasiurus hirsutus. В наиболее глубоко врезанных долинах встречаются крупные кусты ретамы (Retama retam), а изредка и деревья (Acacia raddiana, A. tortilis). Оба упомянутых вида акаций, а также фисташка (Pictacia therebinthus) широко распространены в наиболее увлажненных долинах по всей Северной Сахаре. Местами произрастают сообщества с господством кустарничка зиллы (Zilla spinosa). Кроме упомянутых многолетних видов растений, повсюду присутствуют эфемеры (Trigonella stellata, Mesembryanthemum forskalei, Schismus calycinus, Plantago ovata, Zygophyllum simplex, Matthiola livida, Malva parviflora). В описываемом районе годовое количество осадков не превышает 30 мм, поэтому и общее проективное покрытие растительности на плакорах составляет не более 5%.
Умеренноширотные пустыни Средней Азии (Каракумы, Кызылкум), Ирана и Афганистана принадлежат к тому же седьмому типу, однако отличаются несколько большой увлажненностью за счет прохладных и дождливых зимы и весны. Юг Средней Азии и Восточная Сахара имеют всего лишь около 10% общих видов высших растений, однако на родовом уровне наблюдается очень большое сходство. Обширные площади региона заняты песчаными пустынями. Зимой и ранней весной обильно развиваются эфемеры и эфемероиды (Papaver pavoninum, Senecio, Malcolmia, Carex physodes. Rheum turkestanicum. Ferula oopoda и многие другие). На бугристых песках доминируют кусты белого саксаула (Haloxylon persicum), а в долинообразных понижениях с грунтовым засолением образуется довольно густой «лес» из крупных, высотой до 5 м, кустов черного саксаула (Haloxylon aphyllum). По склонам барханно-грядовых подвижных песков произрастают отдельные кусты кандымов (Calligonum ssp.), эфедры и песчаной «акации» (Ammodendron ssp.). Многолетние травы представлены прежде всего видами Aristida и Dantonia. Самый распространенный вид аристиды - селин (A. karelini). В сильно засоленных понижениях вместо черного саксаула в изобилии растут другие маревые (Salicornia, Halocnemum, Halostachys).
В каменистых гипсовых пустынях также встречаются суккулентные галофиты (Salsola), но многочисленнее колючие и прутовидные формы (Cousinia, Atraphaxis, Gaillonia, Noaea spinosissima, Statice, Reaoumuria). Гипсовые пустыни более богаты и разнообразны в предгорных участках со сложным рельефом (выходы неогеновых пестроцветов и палеогеновых красноцветов). На плакорных выровненных участках эти пустыни обычно имеют обедненный состав и менее обильный растительный покров.
Низкие адыры (плоские предгорья) гор юга Средней Азии, Ирана и Афганистана занимают аридные сообщества особого типа, их часто называют низкотравными полусаваннами, эфемеровыми пустынями, предгорными полупустынями. Несомненно, эфемеровые сообщества на лёссовых отложениях представляют собой переходный вариант между ирано-туранскими пустынями и крайними засушливыми и континентальными вариантами сообществ средиземноморского типа (шибляк, включая аридные редколесья из фисташки и миндаля). В зимне-весеннее время эфемеровые сообщества производят впечатление сообществ чисто мезофильного дуговидного типа. Густой травяной покров формируется в основном из луковичного мятлика (Poa bulbosa), осоки (Carex pachystylis), многочисленных эфемеров (крестоцветные, маковые, сложноцветные, злаки, гвоздичные). Весьма характерны такие геофиты, как тюльпаны, гусиные луки, ферулы, лютики, скорцонеры, герани. Однако с середины мая травяной покров быстро высыхает, желтеет и местность приобретает пустынный облик, сохраняющийся вплоть до зимних дождей.
Запасы фитомассы и величины продукции в разных аридных экосистемах чрезвычайно сильно варьируют - от почти нулевых значений до сравнительно больших величин. Так, в саксаульниках ирано-туранских пустынь, редколесьях из акаций, прозописов, некоторых тропических колючих зарослях запасы сухой фитомассы достигают 50 т/га. В бездревесных сообществах с хорошо развитым травяным покровом (эфемеровые сообщества, некоторые северные полупустыни) эти параметры колеблются в пределах 5-15 т/га. На больших пространствах Северной и Южной Сахары, в пустынях Центральной Азии, Северной Америки фоновые значения запасов сухой фитомассы обычно составляют 2-5 т/га. В наиболее безжизненных пустынях первого и второго типа запасы фитомассы обычно не превышают нескольких десятков килограммов на 1 га.
В большинстве типов аридных экосистем с хорошо развитым древесным или кустарниковым ярусом первичная продукция составляет 6-8 т/га в год. В этих же пределах колеблется продуктивность эфемеровых сообществ лёссовых предгорий Средней Азии, Ирана и Афганистана, а в северных полупустынях эти величины составляют 1-3 т/га в год. В большинстве пустынных экосистем первичная продукция не превышает 1 т/га в год, а в экосистемах первого и второго типов вообще приближается к нулю.
Гетеротрофы первого порядка
Мозаичное и разреженное распределение фотоавтотрофов ведет к такому же неравномерному распределению гетеротрофов первого порядка - потребителей отмершей и живой фитомассы. Из них сапротрофы приурочены к подкроновым участкам древесно-кустарниковых растений с запасами подстилки и к почвенной ризосфере (почве, пронизанной корнями). Именно в ризосфере сосредоточены почти все микроорганизмы, причем бактерии занимают преобладающее положение по отношению к микромицетам. Беспозвоночные животные-сапрофаги представлены сильно обедненными группировками, которые, однако, объединяют много специфических пустынных форм.
Из представителей почвенной мезофауны в большинстве аридных экосистем тропических и субтропических широт ведущее положение занимают термиты. Эти животные ведут зачастую крайне скрытный образ жизни и не сооружают столь заметных жилищ-термитников, как в саваннах, зато их ходами бывают пронизаны глубокие слои грунта (рис. 31). В пустынях с периодическим увлажнением основная деятельность термитов проявляется сразу после окончания периода дождей, когда эти насекомые усиленно потребляют свежеотмершие стебли и опад пустынных растений. В пустынях Африки и Евразии основная часть термитов принадлежит к роду Acanthotermes.
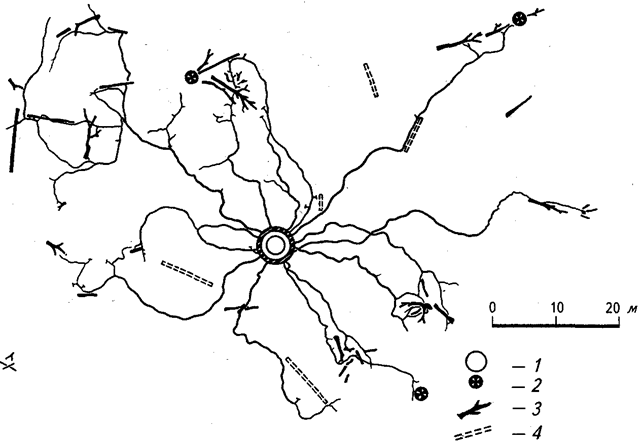
Рис. 31. Система подземных ходов термита Coptotermes lacteus (по К. Ли и Т. Буду, 1975):
1 — центральная надземная постройка термитов; 2 — живые деревья;
3 — валежник; 4 — валежник, переработанный термитами
В пустынных акациевых редколесьях малга-скраб наиболее обычны роды Tumulitermes, Nasutitermes, Coptotermes.
В почвенной толще и в подстилке развиваются личинки многочисленных видов жуков из семейства чернотелок (Tenebrionidae); имаго также потребляют преимущественно растительную пищу в виде сухого опада, семян , молодых зеленых проростков (в сезон дождей). Среди потребителей корней - личинки жуков многих семейств (хрущи, златки, усачи), а также других групп насекомых, в частности бабочек-совок.
Многие пустынные гетеротрофы первого порядка сооружают вертикальные и горизонтальные подземные ходы, перемешивая почву, обогащая ее гумусовыми веществами своих копролитов. Это мокрицы-гемилепистусы, различные тараканы (виды Polyphaga и др.), жуки-навозники (например, Gymnopleurus).
Чрезвычайно характерной группой фитофагов в пустынях на всех материках являются муравьи-жнецы, потребляющие семена (Messor и т. п.). Из беспозвоночных животных, поедающих живую растительную массу, в поверхностных ярусах главное место принадлежит разнообразнейшим саранчовым.
Позвоночные фитофаги представлены в первую очередь грызунами. В разных регионах эти животные зачастую занимают разное систематическое положение (песчанки, крысы, хомяки, гоферовые, гетеромиины, тушканчики, беличьи, свинковые), однако все они объединяются в сравнительно небольшое число функциональных групп. Это наиболее массовые виды - зеленояды, всеядные формы, роющие грызуны, потребляющие подземные части геофитов, семенояды. Многие впадают в спячку на время длительной засухи или морозного периода (во внетропических пустынях) - это суслики, тушканчики. Некоторые виды из разных систематических групп обладают жировыми депо, которые часто локализованы в хвосте (жирнохвостые тушканчики, жирнохвостые песчанки Южной Африки и т. п.).
Более крупные травоядные представлены копытными, которых в Австралии заменяют крупные сумчатые (кенгуру и валлаби). Особенно богаты копытными аридные территории Старого Света. Это различные газели и другие антилопы, дикие ослы Африки, куланы Азии, дикая
Дата добавления: 2019-12-09; просмотров: 1185;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
