Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ

| Сетчатка | ДЗрЯ | Мотонейрон | |||||
| i i Т | |||||||
| Скорость движения глаза | ПмРФМ | ||||||

|
 Сохранение скорости
Сохранение скорости
 Сетчатка
Сетчатка
ЯДЗрТ
РЯКМ
| ♦ | ||||
| вя | ПмРФМ | Мотонейрон | ||


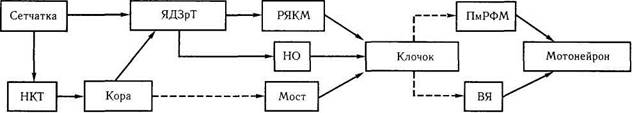
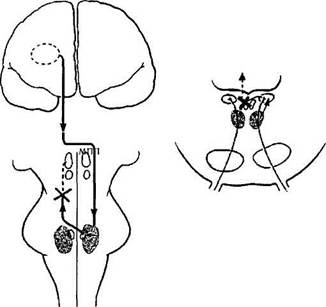
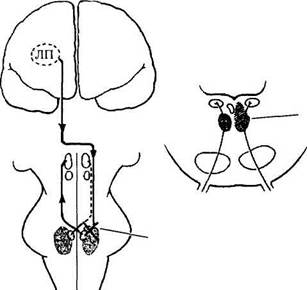
 Рис. 4.4.28. Гипотетические схемы возможных трактов оптокинетической системы (объяснение в тексте):
Рис. 4.4.28. Гипотетические схемы возможных трактов оптокинетической системы (объяснение в тексте):
НКТ — наружное коленчатое тело; ЯДЗрТ — ядра дополнительной системы зрительного тракта; ПмРФМ — парамедианная ретикулярная формация моста; ВЯ — ядра вестибулярной системы; НО — нижняя олива; РЯКМ — ретикулярные ядра крыши моста
кулярном слое. Аксоны этих клеток оканчиваются на теле и дендритах нейронов Пуркине [530].
|
| 18 |
В кору мозжечка входят два типа двигательных волокон. Лазящие (лиановые) волокна проходят через зернистый слой и заканчиваются на дендритах клеток Пуркине. Тела клеток, от которых отходят эти волокна, лежат в ниж-
ней оливе. Все остальные афферентные пути мозжечка представлены мшистыми волокнами, оканчивающимися на клетках-зернах. Каждое мшистое волокно отдает множество коллатера-лей и одно такое волокно иннервирует множество клеток коры мозжечка.
В мозжечок входят и из него выходят крупные пучки нервных волокон (рис. 4.4.30). От ядер моста идут волокна в кору противоположной стороны мозжечка. Этот пучок афферентных волокон называется средней ножкой мозжечка (pedunculus cerebellaris medialis). Афферентные волокна образуют и веревчатое тело, т. е. нижнюю ножку мозжечка (pedunculus cerebellaris inferior). По ней из спинного мозга
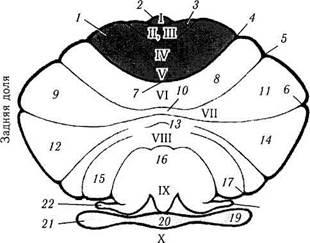
 Рис. 4.4.29. Вентральная поверхность мозжечка после
Рис. 4.4.29. Вентральная поверхность мозжечка после
удаления моста и продолговатого мозга (по Strother,
Salamon, 1977):
I — передняя долька; 2— язычок; 3— центральная долька; 4 — первичная щель; 5 — задне-верхняя щель; 6 — горизонтальная щель; 7— скат; 8— задняя четырехугольня долька; 9 — верхняя полулунная долька; 10— извилина червя; // — ножка I; 12— нижняя полулунная долька; 13— бугорок червя; 14 — ножка II; 15 — двубрюшная долька; 16 — язычок; 17 — препирами-дальная щель; 18 — заднелатеральная щель; 19 — клочок; 20 — узелок; 21 — флоккулонодулярная долька: 22 — парафлоккулус
Движения глаз
 поступает информация о положении конечностей и о состоянии тонуса мышц.
поступает информация о положении конечностей и о состоянии тонуса мышц.
Массивный пучок волокон входит также в контрлатеральное веревчатое тело, а затем в ядра нижней оливы (п. olivaris) продолговатого мозга. Другие волокна в составе нижней ножки мозжечка принадлежат клеткам ретикулярной формации.
Верхняя ножка мозжечка (pedunculus cere-bellaris superior) состоит в основном из эфферентных волокон, проводящих импульсы из мозжечка к образованиям ствола мозга, промежуточного мозга и к полушариям головного мозга (рис. 4.4.30, в).
Мозжечок контролирует движения путем координации сокращения поперчнополосатой мышечной ткани тела, включая наружные мышцы глаза. Подобное управление требует поступления к мозжечку афферентов от зрительных и вестибулярных ядер, проприорецепторов мышц и сочленений шеи, а также проекции эффе-рентов на надъядерные зоны, обеспечивающие движения глаза (плавное слежение и саккады).
Наиболее важными связями являются связи с отдельными структурами мозжечка, приведенные ниже.
Клочково-узелковая доля (lobus flocculo-nodularis) (см. рис. 4.4.29).
Почти все пути от вестибулярной системы оканчиваются в узелке (nodulus) и клочке (flocculus). Давно известен передний спинномоз-жечковой путь (tractus spinocerebellaris anterior) (путь Говерса). Но кроме него известны еще, по крайней мере, 10 путей. Остальные пути представляют собой спинно-оливные пути (tractus spinoolivaris), переключающиеся в оливах на нейроны, посылающие лиановидные волокна к коре мозжечка.
Все пути оканчиваются главным образом в палео- или спиноцеребеллуме. К старому мозжечку идут также соматосенсорные афферентные волокна от головы и зрительные и слуховые афференты.
Зрительная информация достигает клочка через связи, вовлекающие сетчатку, претек-тальную область противоположной стороны, нижнее ядро оливы [530].
Проекции зрительной коры на ядра моста образуют синапсы в каждом околоклочке (para-flocculus) (мшистые волокна) [85].
Ядро зрительного тракта и некоторых нейронов клочка кодируют информацию о направлении и скорости движения цели [117, 513], соответствии движения тела движениям глаза при слежении за целью («сила обратной связи») [203].
Вестибулоцеребеллум модулирует плавное слежение и вестибуло-глазной рефлекс [96, 201, 337, 338]. Дорзальные повреждения вести-булоцеребеллума у человека приводят к потере координации движений глаза с появлением «промахивающихся саккад».
Червь (Vermis). Дорзальный червь, особенно дольки V, VI и VII, и соседнее ядро шатра участвуют в саккадических движениях. Об этом свидетельствует то, что клетки Пуркине червя возбуждаются за 25 мс перед саккадами [343] или во время саккад [242, 292, 297, 530]. Стимуляция этой зоны приводит к содружественным саккадическим движениям.
Афферентные пути состоят из проекций на червь вестибулярных ядер и основных ядер моста. Ядра моста, в свою очередь, получают зрительные афференты от верхних бугорков четверохолмия, наружного коленчатого тела и стриарной коры. Имеются также шейные и глазные проприоцептивные афферентные волокна.
Эфферентные пути косвенно проецируются на глазодвигательные ядра через вестибулярные связи [574].
Полушария мозжечка (hemispherium cere-belli). Проекции полушарий мозжечка складываются из волокон, оставляющих зубчатые ядра и через соединительное плечо достигающих глазодвигательных ядер. Проекция на ядра глазодвигательного нерва происходит непосредственно или после переключения в ретикулярном ядре моста (nucleus reticularis tegmenti pontis) [96, 102, 201, 453], которое лежит в парамедианной ретикулярной формации моста с вентральной стороны [240, 350]. О роли полушарий в движении глаз свидетельствует хотя бы тот факт, что стимуляция полушарий может приводить к саккадическим движениям глаза или плавному слежению [453].
Таким образом, нарушения функции мозжечка довольно разнообразны в своих клинических проявлениях. Диффузные паренхиматозные поражения возникают вследствие вирусного энцефалита, абсцессов, развития опухоли (медулло-бластома, астроцитома), а также наследственных заболеваний типа гемангиобластомы, болезни Гиппель—Линдау (von Hippel—Lindau), мозжечковой дегенерации.
Повреждение передних частей червя приводит в атаксии и затруднению взгляда кверху. Повреждение срединных структур мозжечка сопровождается гипотонией мышц разгибателей конечностей со стороны поражения.
После разрушения мозжечка возможно и развитие повышенного мышечного тонуса, но на короткое время. Это связано с тем, что ядра мозжечка обычно проявляют тормозное влияние на латеральное вестибулярное ядро (nucleus vestibularis lateralis; Дейтерса), которое, в свою очередь, возбуждают альфа-мотонейроны мышц конечностей.
Боковые полушария мозжечка влияют на тонус мышц флексоров через верхнюю ножку мозжечка (соединительное плечо), красное ядро противоположной стороны и красноядер-но-спинномозговой путь с этой же стороны. Поскольку имеется двойной перекрест перечне-
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
ленных путей (за исключением тракта красного ядра), повреждение полушарий мозжечка приводит к гипотонии со стороны поражения. В то же время повреждение красного ядра сопровождается гипотонией противоположной стороны.
Офтальмолог должен помнить и о так называемом глазном миоклоническом треугольнике, образованном полушарием мозжечка, красными ядрами и нижним ядром оливы моста. Эти структуры связаны между собой мозжечковыми ножками и центральными покрышечными путями. Поражение этой области (чаще инфаркт в области моста) приводит к развитию глазной и небной миоклонии.
Нарушение горизонтальных движений глаз.Как показано выше, центром надъядер-ного контроля содружественных движений глаз является парамедианная ретикулярная формация моста. Именно на нее проецируются верхние бугорки четверохолмия, вестибулярные ядра, зрительное поле лобной доли, ядро медиального продольного пучка, мозжечок и nucl. perihypoglossal. Отводящие волокна от этого центра идут к нейронам ипсилатерально-го ядра отводящего нерва, образуют синапсы с мотонейронами и межъядерными (вставочными) нейронами. Аксоны вставочных нейронов перекрещиваются в мосту, формируя медиальный продольный пучок, переходят на противоположную сторону и подходят к ядрам глазодвигательного нерва, иннервирующим внутреннюю прямую мышцу глаза (см. выше).
Нарушение взгляда возможно при повреждении путей на различных уровнях (рис. 4.4.31— 4.4.33). Так,повреждение лобного глазного поля (поле 8) нарушает возможность волевого
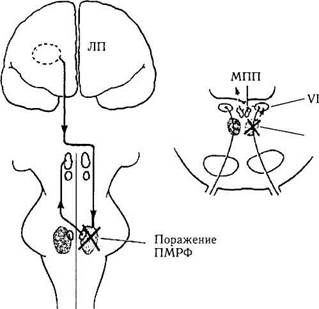
поворота глаз влево. Повреждение области ва-ролиева моста, определяющей горизонтальные движения глаза («центр горизонтального взора»), приводит к параличу наружной прямой мышцы с этой же стороны и контрлатеральному параличу внутренней прямой мышцы. При этом нарушается горизонтальное смещение взора в сторону, соответствующую стороне поражения. Например, если поражена парамедианная ретикулярная формация моста в левой половине мозга, то при попытке больного смотреть прямо оба глаза несколько смещаются влево от средней линии, а полное смещение глаз влево ограничено (рис. 4.4.31). Такие больные также не способны долго смотреть влево или прослеживать глазами объекты, движущиеся в левую сторону поля зрения. При этом не наблюдается атрофии наружной прямой мышцы и диплопии. В тех случаях, когда нарушение в области варолиева моста обширное, с вовлечением кортикоспинальных волокон с этой же стороны, глаза будут смещены в сторону развившейся гемиплегии.
Повреждение левого (правого) ядра отводящего нерва приводит к атрофии левой (правой) наружной прямой мышцы и неспособности больного повернуть глаза кнаружи в левую сторону. Поскольку «мелкие» нейроны левого ядра отводящего нерва погибли, восходящие волокна к контрлатеральному правому ядру и, естественно, волокна, генерирующие внутреннюю прямую мышцу, отсутствуют. В результате этого больной неспособен повернуть правый глаз кнутри, когда он смотрит налево. Необходимо отметить, что атрофии правой внутренней прямой мышцы нет. Возникает вопрос — почему? Этого не наступает по той причине, что нейро-

|
|

|

|
|
| ПМРФ |
Прав.Лев.
О

<Ш> 1 I
\
Вверх
Вниз
Вправо
Влево
Рис. 4.4.31. Нарушение горизонтального взгляда при поражении парамедианной ретикулярной формации (ПМРФ)
Авижения глаз

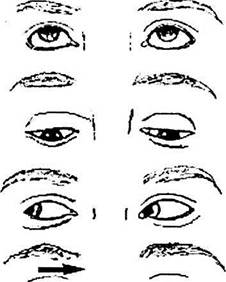
|
|
| f,- |
Прав. Лев.
<ffi> } (О
Вверх
Вниз
Вправо
Влево
Рис. 4.4.32. Нарушение горизонтального взгляда при поражении медиального продольного пучка справа
ны, иннервирующие эту мышцу, не погибают. Исчезает только проекция тракта на них. Об их функции свидетельствует то, что нейроны возбуждаются во время горизонтальных сакка-дических движений влево, при конвергенции. В последнем случае нейроны правой внутренней прямой мышцы глаза получают информацию от «центра конвергенции», расположенного ростральней комплекса глазодвигательных ядер. При поражении ядра отводящего нерва диплопия не развивается.
Несколько иная картина характерна для повреждения отводящего нерва (наиболее часто в пещеристой пазухе). Подобные повреждения приводят к атрофии наружной прямой мышцы глаза со стороны повреждения. По этой причине внутренняя прямая мышца приводит глаз кнутри и развивается диплопия.
Повреждение правого медиального продольного пучка приводит к недостаточности приведения правого глаза и нистагму отведенного глаза (рис. 4.4.32). Нистагм отведенного глаза иницируется тенденцией отведенного глаза к отклонению в назальную сторону, поскольку внутренняя прямая мышца недостаточно инги-бирована в течение абдукции. Это состояние называется межъядерной офтальмоплегией.
Необходимо отметить, что односторонняя межъядерная офтальмоплегия возникает при поражениях, локализованных более кпереди. Это связано с тем, что медиальные продольные пучки разделены в большей степени в среднем мозге и в меньшей степени в области моста (медиальный продольный пучок удобно описывать в виде буквы «V», широкая часть которого располагается в области среднего мозга и узкая в области моста). Встречается она при на-
рушении кровообращения в участке мозга, кро-воснабжаемом базилярной артерией или ее перфорирующими ветвями.
При передней межъядерной офтальмоплегии описаны случаи дополнительного вовлечения покрышки среднего мозга. При этом нарушается конвергенция. У этих больных при содружественном взгляде или конвергенции не сокращаются внутренние прямые мышцы глаза.
Двусторонняя межъядерная офтальмоплегия развивается при поражении более задних отделов и почти всегда встречается при рассеянном склерозе. Сводится она к развитию двустороннего содружественного паралича взгляда (паралич внутренних прямых мышц), сопровождающегося нистагмом отведенных глаз. Двустороннюю межъядерную офтальмоплегию можно отдифференцировать от двустороннего паралича внутренней прямой мышцы, поскольку при стимуляции «конвергентно-аккомодацион-но-зрачкового» рефлекса глаза будут отведены.
Двустороннее поражение медиального продольного пучка сопровождается негоризонтальным нистагмом вертикального или ротационного типов. Это связано с тем, что медиальный продольный пучок несет волокна, обеспечивающие движения глаз в вертикальной плоскости. Медиальный продольный пучок может быть вовлечен в вертикальный нистагм.
Одновременное поражение ретикулярной формации и медиального продольного пучка приводит к нарушению взора, особенности которого проиллюстрированы рис. 4.4.33.
При поражении части медиального продольного пучка, расположенной кзади от ядра отводящего нерва, развиваются многочисленные типы нистагма.
Глава 4.ГОЛОВНОЙ МОЗГ И ГЛАЗ


|
|
| Поражение МПП + ПМРФ |
| Поражение МПП + ПМРФ |
Прав. Лев.
Вверх
Вниз
Вправо
Влево
Рис. 4.4.33. Нарушение горизонтального взгляда при поражении медиального продольного пучка и парамедианной
ретикулярной формации
В заключение необходимо упомянуть и о клиническом проявлении поражения вестибулярного нерва. Например, при повреждении левого вестибулярного нерва начинает контролировать движения глаз правый вестибулярный нерв. Этот дисбаланс приводит к тому, что оба глаза начинают медленно передвигаться влево (правый вестибулярный нерв возбуждает правые вестибулярные ядра, которые, в свою очередь, возбуждают левую парамедианную ретикулярную формацию, а затем и левое ядро отводящего нерва, поворачивающего оба глаза влево). Когда оба глаза максимально отведены влево, они очень быстро поворачиваются вправо. Механизм этого явления пока неясен. Затем глаза медленно передвигаются снова влево, и этот цикл повторяется.
Нарушение вертикальных движений глаз.Центром надъядерного контроля вертикальных движений глаза, также как и при горизонтальных движениях, является парамедианная ретикулярная формация моста. В координацию вертикальных движений также вовлечены интер-стициальное ядро Кахаля, ядро Даркшевича, задняя спайка и ростральное интерстици-
альное ядро медиального продольного пучка [178, 179, 225, 240, 572, 573].
Паралич взгляда в вертикальной плоскости обычно является следствием нарушений в области диенцефало-мезенцефалического соединения. Примером тому могут служить паркинсонизм и прогрессивная надъядерная офтальмоплегия, при которых одновременно нарушен взгляд кверху и книзу.
Паралич взгляда книзу обычно развивается при двусторонних ростральных нарушениях кровообращения, вовлекающих медиальную часть зрительного бугра, fasciculus retroflexus, ядро поводка (п. habenulae) и часть красного ядра (nucleus ruber). Паралич взгляда книзу возникает и при повреждении задней спайки (часто с ретракцией леватора верхнего века, параличом конвергенции и нарушением зрачкового рефлекса), что наиболее часто бывает при возникновении опухоли шишковидной железы или грыжи ствола мозга. В табл. 4.4.7 и 4.4.8 приведены сравнительные характеристики и особенности паралича взгляда книзу и кверху в зависимости от уровня поражения мозга.
Таблица 4.4.7. Сравнительная характеристика вертикального и горизонтального паралича
| Вертикальный | Горизонтальный | |
| Локализация «центра взгляда» | Мезенцефалон | Мост |
| Частота | Нечасто | Обычно |
| Встречаемость полного паралича | Редко | Редко |
| Направление пареза взгляда | Вверх—вниз | Вправо—влево |
| Сопутствующие нарушения | Экстрапирамидные нарушения | Гемипарезы |
| Нарушения чувствительности | Нарушения чувствительности |
движения глаз
 Таблица 4.4.8. Сравнительная характеристика паралича взгляда вверх и вниз
Таблица 4.4.8. Сравнительная характеристика паралича взгляда вверх и вниз
| Паралич взгляда вверх | Паралич взгляда вниз | |
| Частота поражения | Часто | Редко |
| Причина поражения | Опухоль шишковидной железы Инфаркт Гидроцефалия Опухоли мозга | Инфаркт Энцефалопатия Субарохноидальное кровоизлияние Опухоли мозга |
| Локализация поражения | Дорзальная часть среднего мозга Область сильвиевого водопровода | Вентральная часть мезодиенцефалическо-го соединения Дорзомедиальная часть красного ядра Вентральная часть переакведуктального серого вещества |
| Зрачок | Диссоциация конвергентно-аккомода-ционно-зрачкового рефлекса Обычно расширен | Нормальный или расширен |
| Опущение века | + | — |
| Утеря феномена Белла | + | — |
| Сторона поражения | Обычно, но не всегда, двухстороннее | Всегда двухстороннее |
Повреждения в области моста или покрышки среднего мозга, вовлечение надъядерных волокон, направляющихся только к одному глазу, могут вызвать несодружественное вертикальное положение глаз в покое. Подобное состояние должно быть дифференцировано от состояния, развивающегося при повреждении ствола мозга. В последнем случае глаза отклоняются горизонтально и вертикально.
Могут также развиваться надъядерные тонические и фазовые вертикальные отклонения глаз. Тонические отклонения встречаются и в норме в грудном возрасте. У взрослых отклонение глаз книзу иногда наступает при инфаркте в области зрительного бугра, метаболической энцефалопатии или гидроцефалии. При этом иногда веко опущено.
Двустороннее поражение ретикулярной формации среднего мозга вызывает паралич вертикальных движений глаз, в результате чего становится невозможным произвольно выполнить поворот глаз вверх или вниз.
Движенияглаз во сне.Признано, что имеется два типа движений во сне — быстрые и медленные. Медленный тип встречается большую часть времени, но он прерывается периодами быстрых движений глаз. При этом содружественные движения глаз наблюдаются в любом направлении. Они зависят от путей, простирающихся от ретикулярной формации варо-лиева моста через наружные коленчатые тела до затылочной коры. Эти движения глаз легко можно наблюдать и через закрытые веки.
Движения глаз и восприятие движения.В заключение раздела целесообразно остановиться на вопросах, связанных не с анатомией, а скорее, с нейрофизиологией и нейропсихологией. Связано это с тем, что исследования анатомического, физиологического характера да-
леко не полностью объясняют значение движений глаз в процессе восприятия и анализа изображения. Существует много непонятных до сих пор явлений, которые, в какой-то мере, можно объяснить, используя знания, полученные в области нейропсихологии.
Первоначально необходимо отметить то, что основной функцией движений глаз, описанных выше, является поиск зрительной цели и проекция ее на центральную ямку. Даже после выполнения этой функции для восприятия изображения необходимы движения, называемые микросаккадами («физиологический нистагм»). Это очень быстрые и мелкие движения, при которых глаз перемещается примерно на 20° угловых секунд 30—60 раз в секунду. Именно благодаря этим движениям и происходит в сетчатой оболочке формирование нервного импульса. При полном прекращении движений глаза, что можно достичь в экспериментальных условиях полной акинезией наружных мышц глаза или специальными оптическими системами, восприятие изображения исчезает [5]. Микросаккады реализуют основное условие возбуждение рецептивных полей сетчатой оболочки, являющихся детекторами распознавания признаков изображения (линии, края, окружности, цвета и др.). Когда изображение становится неподвижным относительно сетчатки, рецепторы перестают посылать сигналы в центральную нервную систему, они перестают функционировать и зрительный образ исчезает.
Обращает на себя внимание и следующий факт. Давно было отмечено, что если изображение объекта перемещается по сетчатке неподвижного глаза, то воспринимается движение этого объекта. В то же время если глаз следит за движущимся объектом, то его изображение остается в том же месте на сетчатке,
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 но при этом движение объекта все же воспринимается. Вместе с тем при произвольном изменении направления взора неподвижные объекты не воспринимаются как движущиеся, хотя в действительности изображения этих объектов при саккадических движениях смещаются по сетчатке. Наоборот, если слегка надавливать пальцем на глаз, то такое пассивное движение глаз вызывает смещение изображения на сетчатке, которое интерпретируется как движение окружающих предметов. Более того, при нистагме, вызванном стимуляцией вестибулярной системы или алкогольной интоксикацией, смещение изображения на сетчатке во время медленной фазы нистагма воспринимается как движение окружающих предметов.
но при этом движение объекта все же воспринимается. Вместе с тем при произвольном изменении направления взора неподвижные объекты не воспринимаются как движущиеся, хотя в действительности изображения этих объектов при саккадических движениях смещаются по сетчатке. Наоборот, если слегка надавливать пальцем на глаз, то такое пассивное движение глаз вызывает смещение изображения на сетчатке, которое интерпретируется как движение окружающих предметов. Более того, при нистагме, вызванном стимуляцией вестибулярной системы или алкогольной интоксикацией, смещение изображения на сетчатке во время медленной фазы нистагма воспринимается как движение окружающих предметов.
На основании этих наблюдений Гельмгольц заключил, что обработка зрительной информации в мозге должна включать операцию сравнения эфферентных команд на движение глаз и головы с афферентными зрительными сигналами, поступающими от сетчатки. Данный механизм был назван Хольстом и Миттельштадтом принципом «реафферентации». При этом команды из центральных отделов мозга на изменение направления взора передаются через области, управляющие взором, на ядра, управляющие сокращением мышц. Результатом этого движения будет смещение на сетчатке изображений всех неподвижных предметов. Однако в этом случае никакого смещения объектов не воспринимается, поскольку в центральной нервной системе сохраняется копия эфферентных сигналов, задающих движения глаз («эфферентная копия»). Путем сравнения этой копии с сигналами, отражающими смещение изображения на сетчатке, может быть установлена причина такого смещения. Если же глаз следит за небольшим равномерно движущимся объектом, то смещение изображения на сетчатке исключительно мало, а движение его воспринимается. Считается, что в этом случае восприятие движения осуществляется за счет сравнения сигналов от сетчатки с эфферентной копией движения.
Теория «эфферентной копии» нашла широкое распространение, тем более что на основе ее использования возможно количественно оценить физиологические явления. Ясно, что копия локализуется в коре мозга, но точно место ее расположения не определено. Кора четко фиксирует расчетное положение глаза в определенный момент, полученное в результате афферентных сигналов, идущих от каждой пары мышц антогонистов. «Эфферентная копия» используется для вычисления положения глаз до начала их движения и рефиксируется после саккады.
«Эфферентная копия» поставляет информацию, посылаемую каждому глазу так, что «ней-ральный интегратор» горизонтального взора создает четыре «эфферентные копии» от четырех горизонтальных наружных мышц глаза.
Эфферентные сигналы могут быть противопоставлены афферентным сигналам, получаемым от системы контроля положения глаза.
4.5. ВЕГЕТАТИВНАЯ (АВТОНОМНАЯ) ИННЕРВАЦИЯ ГААЗА
4.5.1. Вегетативная система
и ее контроль центральной нервной системой
Вегетативная нервная система, иннервируя гладкую мускулатуру всех органов, кровеносных сосудов, сердце и железы, отвечает за регуляцию внутренней среды организма [7, 9, 578]. Для офтальмолога наиболее важным является то, что она обеспечивает зрачковый рефлекс, аккомодацию, секреторную функцию слезной железы [412]. Под ее контролем находится внутриглазное давление, функции различных структур глаза и глазницы [202].
Вегетативная (автономная) нервная система получила свое название в связи с тем, что ранее предполагали полное отсутствие контроля над ней со стороны коры головного мозга, поскольку функционирует она даже при нарушении связи между спинным и головным мозгом. Именно этим вегетативная нервная система отличается от произвольной, сознательно управляемой, соматической системы.
Наиболее высокими уровнями контроля деятельности вегетативной нервной системы являются стволовая часть мозга, гипоталамус и лимбическая система. Эти структуры вовлечены в большинство жизненно важных «бессознательных» функций по обработке информации, поступающей от органов и тканей организма, и управлению их деятельностью. В свою очередь, стволовая часть мозга, гипоталамус и лимбическая система находятся под волевым контролем коры головного мозга. Таким образом, понятие автономии вегетативной нервной системы довольно относительно.
О значении коры головного мозга и нижележащих структур в деятельности вегетативной нервной системы свидетельствует хотя бы такой факт. Стимуляция коры лобной, затылочной долей, а также стимуляция многих областей промежуточного мозга вызывает сужение или расширение зрачка.
Большую роль играет гипоталамус. Описано развитие синдрома Горнера после случайных повреждений гипоталамуса во время проведения стереотаксических операций. Стимуляция каудальной части гипоталамуса и серого вещества ствола приводит к расширению зрачка, в то время как их разрушение приводит к сонливости и сужению зрачка. О роли гипоталамуса в деятельности автономной системы свидетельствует и активация ее при сильном эмоцио-
Вегетативная (автономная) иннервация глаза
 нальном возбуждении. Кроме того, гипоталамус обеспечивает надъядерное торможение зрачкового рефлекса, усиливающееся с возрастом.
нальном возбуждении. Кроме того, гипоталамус обеспечивает надъядерное торможение зрачкового рефлекса, усиливающееся с возрастом.
Вегетативная нервная система существенно отличается от соматической и особенностями структурной организации. В первую очередь, она является двухнейронной системой. Один синапс образуется уже после выхода из центральной нервной системы в ганглиях, а второй синапс сформирован уже в эффекторном органе.
Следующее отличие сводится к тому, что соматическая нервная система формирует синапс (нейромышечный), обладающий достаточно стабильной структурой, в то время как синапсы автономной нервной системы представляют собой довольно разнообразные по строению структуры, диффузно распределенные на эффекторном органе.
В функциональном отношении обращает на себя внимание то, что если при стимуляции соматической нервной системы происходит возбуждение эффекторного органа (мышцы), то при стимуляции вегетативной могут наблюдаться как явления возбуждения, так и торможения.
В своей деятельности автономная нервная система использует большое количество различных типов нейромедиаторов и рецепторов.
Существуют различия и в функциональных проявлениях посттравматической регенерации вегетативных и соматических нервов. После денервации мышцы, иннервируемой автономной нервной системой, снижается мышечный тонус, но не наступает истинного паралича. В последующем нормальный тонус восстанавливается, а также возможно развитие гиперчувствительности мышцы к медиаторам (ацетилхолину для парасимпатической системы, норадрена-лину для симпатической системы). Фармакологические механизмы гиперчувствительности при денервации симпатической и парасимпатической нервной системы различны. В первом случае определяется преюнкциональная, а во втором — постъюнкциональная гиперчувствительность. Преюнкциональная гиперчувствительность связана с утерей способности пресинаптическим аксоном поглощать избыток медиатора, в результате чего существенно увеличивается концентрация норадреналина в области синапса. Постъюнкциональная гиперчувствительность связана со структурно-функциональным изменением самой мышцы. При этом отмечается утеря специфичности рецептора к нейромедиатору.
В структурном отношении периферический отдел вегетативной нервной системы является исключительно эфферентным. Нейроны, расположенные в стволовой части мозга и спинном мозге, и их аксоны, направляющиеся к вегетативным ганглиям, называются преганглионар-ными нейронами. Нейроны, лежащие в вегетативных ганглиях, называются постганглионар-
ными, так как их аксоны выходят из ганглиев и идут к исполнительным органам (рис. 4.5.1).
Аксоны преганглионарных нейронов обладают миелиновой оболочкой. По этой причине их еще называют белыми нервными веточками. Аксоны постганглионарных нейронов немиели-низированы (серые веточки), за исключением постганглионарных аксонов, исходящих из ресничного ганглия. Направляясь к исполнительному органу, вегетативные нервы образуют в их стенке густое сплетение.
Как было указано выше, периферическую часть вегетативной нервной системы подразделяют на два отдела — симпатическую и парасимпатическую. Центры этих отделов лежат на разных уровнях центральной нервной системы.
Многие внутренние органы получают как симпатическую, так и парасимпатическую иннервацию. Влияние этих двух отделов часто носит антагонистический характер, а нередко действуют «синергично». В физиологических условиях деятельность органов зависит от преобладания влияния той или иной системы. Основные структурные и функциональные особенности вегетативной иннервации органов и тканей человека представлены на рис. 4.5.1.
Дата добавления: 2016-06-05; просмотров: 2881;
Поиск по сайту
Узнать еще
- I. Синусы твердой оболочки головного мозга.
- II. Анатомия продолговатого мозга, ретикулярная формация
- IV. проводящие пути головного и спинного мозга
- V Патопсихологическое – при нарушении целостности мозга происходит нарушение психической деятельности
- VII. Вены глазницы.
- А Нейропсихологические синдромы поражения задних отделов коры больших полушарий головного мозга.
- Абсцессы головного мозга. Хирургическое лечение
- Альтернирующие синдромы при поражении ствола головного мозга

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
