Структурно-функциональная организация первичной зрительной коры
Глазные доминантные колонки.После выявления основных физиологических закономерностей реакции зрительной коры головного мозга на «световые стимулы» возник вопрос — какова структурная организация коры, обеспечивающая эти физиологические особенности? Решению этих вопросов содействовало применение новых методов исследования. На протяжении последних двух десятилетий нейронную структуру и функцию зрительной коры интенсивно изучали путем маркировки аксонов нейронов ганглиозных клеток сетчатки и наружного коленчатого тела. При исследовании терминальной дегенерации использовали методы радиоаутографии, а ретроградной дегенерации — введение пероксидазы хрена [256—270, 326, 333, 415]. Большинство исследований проведено на кошках и обезьянах. Показано, что многие закономерности организации коры, выявленные у этих животных, распространяются и на человека.
| Рис. 4.2.55. Доминантные колонки зрительной коры |
В настоящее время ясно, что в структурно-функциональном отношении нейроны первичной зрительной коры организованы в так называемые «колоночные единицы» [540] (рис. 4.2.50, 4.2.55, 4.2.56). Анатомическая реальность «колоночной» цитоархитектоники зрительной коры подтверждена методами марки-

Функциональная анатомия зрительной системы

|
| Зрительное поле (Y2) |
| Стриарная кора (VI) |
| Верхние бугорки четверохолмия |
Наружное коленчатое тело
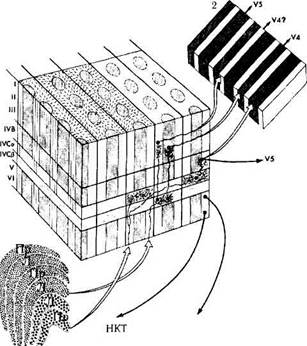
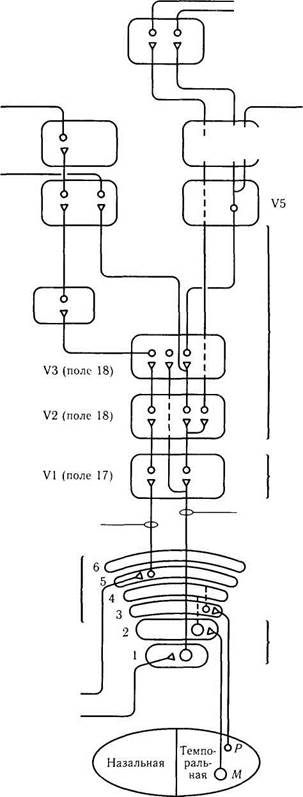
Рис. 4.2.56. Схематическое изображение магно- (М) и парвоцеллюлярных (Р) трактов, идущих от наружного коленчатого тела через поля VI и V2 к полям V4 и V5:
каждый модуль стриарной коры содержит определенное количество глазных доминантных колонок, ориентационных колонок и дающих положительную реакцию при выявлении активности ци-тохромоксидазы «капель» (заштрихованные цилиндры, которые прерываются в слое IVC). Ориентационные колонки (отмечены наклонными линиями на поверхности коры) распространяются через все слои, за исключением слоя IVB-бета. Магноцеллюляр-ный путь проходит через слои IVC-альфа, IVB и направляется к полям коры V2 и V5. Парвоцеллюлярный путь проецируется через слой IVC-бета на слои II и III. Клетки цитохромоксидазных капель проецируются на тонкую темную полосу в поле V2. Клетки, расположенные между капель, проецируются на светлые тонкие полосы V2, а затем, возможно, на V4. Слои V и VI проецируются на верхние бугорки четверохолмия и наружное коленчатое тело соответственно
ровки нейронов индикаторами типа радиоактивного пролина. После введения радиоактивного пролина внутрь глаза он распространяется по направлению наружного коленчатого тела, а затем через синапсы к первичной зрительной коре [259]. При использовании этого метода зрительная кора неравномерно окрашивается в виде «колонок». У человека «колонки» выявляются при определении активности цито-хромоксидазы.
Благодаря использованию подобных методов установлено, что нервные клетки зрительной коры распространяются не только по слоям, параллельным поверхности коры, но также существуют функциональные слои, расположенные перпендикулярно поверхности коры. Подобные структурные образования отсутствуют в месте проекции слепого пятна и «назального полумесяца».
Электрофизиологическая регистрация сигнала от нейронов IVC-слоя зрительной коры по-
зволила установить, что информация от наружного коленчатого тела направляется к смежным «колонкам», относящимся к правому и левому глазам, чередуясь при этом (рис. 4.2.50, 4.2.56). Подобные отношения структурно обеспечивают бинокулярные взаимоотношения на уровне коры. Монокулярно доминирующие «колонки» названы «глазодоминантными колонками». Именно расположением «глазодоминантных колонок» обоих глаз по соседству, а также наличием нейронных связей между ними обеспечивается «перекрывание» точек зрительного пространства, проецирующихся на левое и правое глазное яблоко.
Колоночные комплексы нейронов первичной зрительной коры большего размера, включающие в свой состав «глазодоминантные колонки» левого и правого глаза, а также, описанные ниже «ориентационные колонки», называются «гиперколонками». Площадь среза «гиперколонки» равняется примерно 1 мм2. На каждую «гиперколонку» проецируется определенная область зрительного поля. Причем размер «гиперколонки» зависит от участка сетчатой оболочки, проецирующейся на кору [348, 349, 451, 584, 607]. Наибольшим размером отличаются «гиперколонки», на которые проецируется периферия сетчатки, а наименьшим — маку-лярная область. Правда, необходимо отметить, что полного соответствия проекции сетчатой оболочки на зрительную кору не определяется, что является причиной дальнейших интенсивных исследований с использованием методов электрофизиологии [60]. Колончатой структурой обладают и детекторы цвета. В «цветовой колонке» нейроны реагируют только на монохроматическое излучение в узком диапазоне спектра. Каждая «колонка» детектирует специфическую полосу спектра. «Цветовые колонки» чередуются. «Колонки» с нейронами, селективно выделяющими область 490—500 нм, сменяются «колонками», выделяющими область 610 нм и т. д.
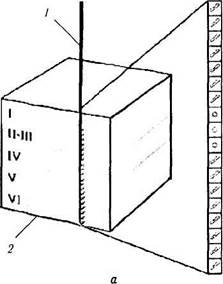
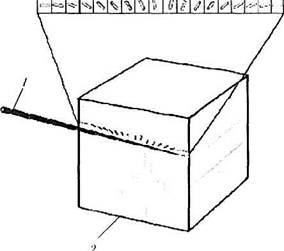
Ориентационные колонки и обработка информации.Помимо «глазодоминантных колонок» в зрительной коре выявлены и другие структурно-функциональные единицы, определяющие возможность определения пространственной ориентации зрительного объекта [65, 155]. Эти структуры были обнаружены благодаря использованию методов электрофизиологии. Впервые наличие ориентационной организации зрительной коры выявили Hubel и Wiesel [262, 263]. Вводя микроэлектрод строго перпендикулярно поверхности первичной зрительной коры, авторы установили, что все нейроны этой области отвечают только при одном условии — наличии определенной ориентации «светового сигнала» в виде щели или грани. Эти данные указывают на то, что эти нейроны сгруппированы в «колонки». Названы они «ориентацион-ными колонками» (рис. 4.2.57).
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ

|
|
Рис. 4.2.57. Схематическое изображение, иллюстрирующее ответ нейрона первичнои зрительной коры при
проникновении микроэлектрода:
а — строго перпендикулярно поверхности коры: б—параллельно поверхности коры (/ —микроэлгкгрод: 2 — «ориентационные
колонки»; I—VI — слои первичной зрительной коры)
После введения микроэлектрода в зрительную кору на определенную глубину коры для достижения максимального ответа рецептивного поля Hubel, Wiesel [270] требовалось изменять ориентацию световой щели, поворачавая ее по часовой стрелке или наоборот. Введение электорода на глубину от 1 до 2 мм обычно достаточно для пересечения «ориентационных колонок», охватывающих 180° зрительного пространства обоих глаз.
Результаты электрофизиологических исследований, свидетельствующие о существовании ориентационных колонок, вскоре были подтверждены многочисленными гистохимическими и физиологическими методами [см. обзор 149]. Благодаря использованию этих методов установлено, что «ориентационные колонки» формируют сложную систему полос, органически взаимодействующих с «глазодоминантными колонками». Две системы колонок взаимодействуют настолько интимно, что любой данный блок корковой ткани, площадь которого равняется нескольким квадратным миллиметрам, содержит полный набор «ориентационных» (0° к 180°) и «глазных доминантных колонок». Такой модуль корковой ткани содержит клетки, чувствительные на все ориентации, представленные в обоих глазных яблоках.
Hubel и Wiesel [270] предполагают, что первичная зрительная кора разделена на сотни модулей, каждый из которых содержит законченную систему колонок для анализа зрительной информации от любой данной части поля зрения. Наибольшее количество подобных мо-
дулей расположено в коре, обеспечивающей центральное зрение.
«Пятна» активности цитохромоксидазы.
Предположение Hubel и Wiesel относительно существования нескольких сотен блоков ткани было подтверждено при определении активности цитохромоксидазы нейронов зрительной коры [593]. Определение активности цитохромоксидазы в зрительной коре показало распределение активности фермента в виде овальных пятен. Подобные пятна обнаруживаются во всех слоях коры, за исключением слоев IVC и IVA [252, 253]. Эти участки были названы «каплей» (рис. 4.2.58). Подобные образования обнаружены также у человека. Каждая «капля» фактически является профилем вертикального цилиндра, проходящего через все слои коры. Эти участки являются местом повышенной метаболической активности первичной зрительной коры и могут быть визуализированы после определения активности не только цитохромоксидазы, но и ряда других ферментов.
У обезьян «капли» активности цитохромоксидазы в коре расположены длинными рядами, разделенными светлыми пространствами, шириной 400 мим. После энуклеации одного глазного яблока «капли» цитохромоксидазы становятся более светлыми и узкими, что отражает снижение метаболической активности нейронов. На «капли» прямо проецируются как пар-во-, так и магноцеллюлярные слои наружного коленчатого тела.
После открытия «капель» были проведены многочисленные физиологические исследова-
Функциональная анатомия зрительной системы


|
Рис. 4 2 58. «Капли» активности цитохромоксида-зы первичной зрительной коры (по Horion, 1984):
активность цитохромоксидазы. формирующая струмчры в виде «капель», распространяется на слои II и III первичной зрительной коры (тангенциальный срез). «Пятна» орисш ированы в виде длинных рядов, расположенных параллельно «доминантным колонкам» IVc слоя. Ряды капель» пересекают границу между VI и V2 полями {маленькие стрелки). Лучше видна граница справа (изогнутая стрелка). Полосы различной плотности поля V2 довольно четко видны справа {короткие толстые стрелки)
ния, направленные на выяснение функций II и III слоев первичной зрительной коры, в которых они преимущественно и располагаются. При этом в пределах «капель» обнаружены группы клеток, отличающиеся «специальными» свойствами рецептивного поля, кардинально отличающиеся от рецептивных полей «комплексных» и «ориентационных клеток», расположенных между «каплями». Клетки в пределах «капель» имеют циркулярную симметричную организацию рецептивного поля и поэтому не относятся к ориентационным. При этом около 70% комплексов нейронов отвечают на цветные стимулы [149, 340, 553, 555]. Большинство этих «цветовых» клеток напоминают четыре типа клеток, описанных в наружном коленчатом теле обезьян. Тем не менее обнаружен и неизвестный ранее тип клетки, обеспечивающий, как предполагают, канал с высокой разрешающей способностью информации относительно цветового контраста.
Структурно-функциональная организация ассоциативной зрительной коры(поля 18, 19). Согласно классическому представлению, первичная зрительная кора выполняет первичный анализ зрительной информации, поступающей от наружного коленчатого тела, а затем передает некоторую «эссенцию», полученную в результате обработки информации, ассоциативным зрительным областям.
Существование многоконтактных зрительных областей коры головного мозга явилось довольно существенным открытием последней четверти двадцатого столетия в области сен-
сорной биологии. Обширные участки коры (более 50% площади поверхности) прямо или косвенно вовлечены в обработку зрительной информации.
Ранее считали, что зрительное восприятие определяется двумя зрительными зависимыми областями. К ним относятся поля 18 и 19. Их называют также вторичной и третичной зрительной корой (рис. 4.2.46, 4.2.59, 4.2.60).

Рис. 4.2.59. Горизонтальный срез через заднюю часть головного мозга, иллюстрирующий гистологические особенности зрительной коры (первичная зрительная кора и ассоциативные области) (по Zeki, 1995):
часть коры, расположенная слева (VI), относится к первичной зрительной коре. Эта часть коры по цитоархитектонике отличается от коры, расположенной справа (престриарная кора). Стрелка указывает место перехода стриарной в престриарную кору. Престриарная кора состоит из нескольких зрительных полей (V2—V5), отличающихся в функциональном отношении
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ

|
| Вид с внутренней стороны |
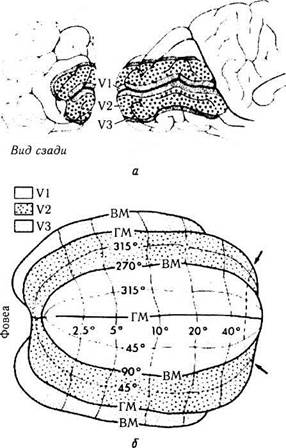
Рис. 4.2.60. Расположение зрительных полей VI, V2 и V3 вдоль шпорной борозды левой затылочной доли мозга (а) и представительство в них зрительных полей, представленных в плоских координатах (б) (по Horton, Hoyt. 1991)
В настоящее время установлено, что в этих процессах, помимо обширных областей затылочной коры, участвуют также височная и теменная кора. Причем каждая из областей выполняет строго определенные функции (см. ниже). Самой важной закономерностью функционирования экстрастриарнои коры является иерархическая и параллельная обработка зрительной информации. Более того, экстра-стриарные области обладают ретинотопически-ми картами зрительного пространства. Известны и стереотопические проекции от одной зрительной области к другой. Эти связи, как правило, двухсторонние.
Первоначально мы охарактеризуем 18-е и 19-е поля.
Вторичная зрительная кора (поле 18) непосредственно прилежит к 17-му полю. Полоска Дженнери в ней отсутствует. На вторичную зрительную кору (кора нижней височной доли; поле 18), т. е. место, определяющее зрительную память, проецируется 17-е поле. Наличие подобной проекции объясняет механизм развития зрительной дезориентации, потери «топографической» памяти, зрительной агнозии и потерю
способности оценить расстояние до объекта при повреждении зрительной коры.
Третичная зрительная кора (поле 19) окружает вторичную кору на медиальной поверхности полушария мозга. Большая ее часть переходит на заднюю теменную долю, а снизу она формирует часть височной доли. При гистологическом исследовании третичная зрительная кора напоминает кору теменной доли, за исключением того, что в V слое отсутствуют большие пирамидные нейроны.
В срезах, окрашенных по Нисслю, эти поля выглядят довольно однородно и иногда даже квалифицированные анатомы не могут найти существенных различий между ними. Последние исследования с использованием электрофизиологических методов и радиоактивных трей-серов выявили в полях 18 и 19, по крайней мере, пять подобластей, отличающихся в функциональном отношении и принимающих участие в обработке зрительной информации. Это поля V2, V3, V3A, V4 и V5 [60, 77, 144] (рис. 4.2.59, 4.2.61, 4.2.62). Существует значительно больше полей, анализирующих зрительную информацию, но расположенных вне полей 18 и 19. К настоящему моменту у обезьян выявлено 25 подобных областей коры и семь ассоциативных областей, обеспечивающих связи с другими известными зрительными областями [322]. Эти области связаны между собой 305 отдельными проводящими путями. 32 зрительные области у обезьян занимают приблизительно 55% площади неокортекса. Это указывает на то, что обработка зрительной информации требует большого количества ткани коры мозга. Первичная зрительная кора в среднем составляет 1200 мм2, или приблизительно 12% неокортекса. Поле V2 несколько меньше и является второй по величине корковой областью. Вместе поля VI и V2 занимают примерно 20% полной площади поверхности неокортекса. Увеличение площади
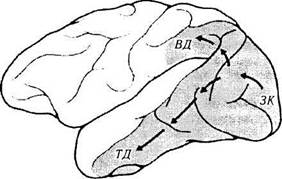
Рис. 4.2.61. Зрительные поля коры головного мозга обезьяны (по Mishkin, 1983):
стрелками указаны два кортикальных зрительных пути, начинающихся из первичной зрительной коры (ЗК). Первый путь направляется вентрально по направлению височной коры (ВД). Второй путь направляется дорзально в кору теменной доли (ТД). Вентральный путь является ключевым в объектном зрении, а дорзальный в пространственном
Функциональная анатомия зрительной системы
■157

|
|
| КОРА |
| Лобное глазное поле (поле 8) |
| Нижняя височная (поля 20, 21) |
| I 1 | Задняя I ' париетальная I I 7 I (поле 7) |
| V4 (поле 18) |
| V3a (поле 18) |
| Престриарная кора |
| Первичная зрительная кора |
| М (магноцеллюлярный тракт) |
| Р (парвоцеллюлярный тракт) |
| Парвоцеллюлярные слои |
| Наружное коленчатое тело |
| СЕТЧАТКА |
Рис. 4.2.62. Схема, иллюстрирующая существование двойственности зрительного пути (по Kandell, Schwartz,
1985):
с одной стороны, зрительный путь характеризуется наличием довольно сложной иерархии структурных образований, идущих от сетчатой оболочки до различных участков коры головного мозга. С другой — существует два параллельных пути передачи зрительной информации — Р- и М-тракты. Парвоцеллюлярный (Я) тракт обеспечивает восприятие деталей, формы и цвета зрительного объекта, в то время как магноцеллюлярный (М) тракт анализирует движение цели
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 ассоциативных областей отражает процессы эволюционного совершенствования анализа зрительной информации. Этот же процесс отражает наличие многочисленных связей ассоциативной коры с другими участками мозга. Как было указано выше, на эти поля проецируется первичная зрительная кора. Благодаря волокнам мозолистого тела происходит соединение 18-го и 19-го полей одного полушария с аналогичными полями противоположного полушария и, таким образом, структурно обеспечивается взаимодействие обоих полушарий мозга. Кроме того, ассоциативные зрительные поля проецируются на кору других долей мозга (лобная, височная, теменная кора) (рис. 4.2.61). Именно последние и обеспечивают высокую степень интеграции и обработки зрительной информации.
ассоциативных областей отражает процессы эволюционного совершенствования анализа зрительной информации. Этот же процесс отражает наличие многочисленных связей ассоциативной коры с другими участками мозга. Как было указано выше, на эти поля проецируется первичная зрительная кора. Благодаря волокнам мозолистого тела происходит соединение 18-го и 19-го полей одного полушария с аналогичными полями противоположного полушария и, таким образом, структурно обеспечивается взаимодействие обоих полушарий мозга. Кроме того, ассоциативные зрительные поля проецируются на кору других долей мозга (лобная, височная, теменная кора) (рис. 4.2.61). Именно последние и обеспечивают высокую степень интеграции и обработки зрительной информации.
В этом разделе мы кратко остановимся только на ассоциативных областях, изученных наиболее полно.
Поля V2 и V3. Границу между полями V2 и V3 стало возможным определить после проведения экспериментов, которые сводились к рассечению волокон мозолистого тела, что приводит к нарушению связи между правым и левым полушариями мозга. Проходящие в мозолистом теле волокна объединяют зрительную кору двух полушарий, проецируя вертикальное сечение поля зрения на поля VI, V2 и V3. Исследование распределения дегенеративно измененных синапсов выявило, что представительство вертикального сечения в поле V2 разделено по линии границы с полем VI (рис. 4.2.59). Представительство горизонтального сечения в пределах V2 поля охватывает с внешней стороны поле V2 по границе, проходящей между полями V2 и V3. Внешняя граница поля V3 представляет вертикальное сечение.
Поля VI, V2 и V3 размещены таким образом, что формируется организованное топографическое представительство зрительных полу-полей в каждой области, в то время как контакт развернут между смежными корковыми областями (рис. 4.2.60). Благодаря этому мозг достигает оптимального взаимоотношения между ретинотопической организацией и необходимостью смежного расположения структурно-функциональных единиц правого и левого глаз. При этом поля V2 и V3 разделяются на дорзальные и вентральные половины, окружающие поле VI. В результате этого нижние и верхние квадранты поля зрения отображены ретинотопически в полях V2 и V3, но прерывисты поперек горизонтального сечения. V3 фактически полностью разделено на отдельные дорзальные и вентральные половины, представляя более низкие и верхние контрлатеральные квадранты поля зрения соответственно (рис. 4.2.60).
Дорзальное и вентральное подполя V3 поля отличаются анатомическими связями и физиологическими свойствами.
Использование гистохимического метода выявления активности цитохромоксидазы позволило установить, что в поле V2 существуют широкие параллельные полосы (рис. 4.2.54) [342]. Этим поле V2 весьма отличается от поля VI, которому свойственны каплеподобные участки активности фермента («капли»). Полосы, характеризующие активность фермента в цитоплазме нейронов V2 поля, имеют различную ширину и интенсивность окрашивания. Различают толстые, тонкие и бледные полосы, отличающиеся особенностями проекции на них первичной зрительной коры и, естественно, функциональными особенностями. Простираются они поперек полной ширины V2 поля от границы полей VI—V2 до границы полей V2—V3.
На тонкие полосы проецируются «капли» первичной зрительной коры. Их нейроны не отличаются ориентационной чувствительностью. Многие из них обладают цветооппонент-ными свойствами. Предполагаемый тракт, завершающийся в этих полосах, имеет следующий вид: нейроны парво- и магноцеллюлярных слоев наружного коленчатого тела —> VI слой, подслой 4Ь —» «капли» —» тонкие полосы.
На «толстые» полосы проецируются участки VI поля, расположенные между «каплями». Нейроны этих полос чувствительны к ориентации «светового стимула», но не чувствительны к их направлению и движению. Половина этих клеток относится к «комплексным» клеткам энд-стопт (end-stopped) типа. Нейроны не реагируют на спектральные характеристики возбуждающего света. Предполагают, что тракт «толстых» полос имеет следующий вид: нейроны парвоцеллюлярных слоев наружного коленчатого тела —» VI слой, подслой 4 СЬ -> —» «интеркапли» —* «толстые» полосы.
«Бледные» полосы получают зрительную информацию от слоя 4В первичной зрительной коры (VI). Клетки отличаются чувствительностью к ориентации зрительного стимула. Часть клеток относится к end-stopped типу. Они не чувствительны к цвету, но регистрируют наличие диспарантности сетчаток. Характеризуется это тем, что стимуляция одного глаза приводит к возникновению потенциала, амплитуда которого значительно меньше, чем при стимуляции одновременно обоих глаз.
Предполагаемый тракт выглядит следующим образом: нейроны магноцеллюлярного слоя наружного коленчатого тела —> 4В слой первичной зрительной коры (VI) —>■ «бледные полосы».
Приведенные выше данные позволяют предполагать, что поле V2 подразделено на ряд под-полей, отличающихся функциональными свойствами. Особенности связей поля V2 с первичной зрительной корой и другими зрительноми областями свидетельствуют о том, что поле V2 определяет достаточно высокий уровень обработки зрительной информации.
Функциональная анатомия зрительной системы
 В последнее время особое внимание нейрофизиологами обращено на поле V3A. Именно это поле обладает многочисленными связями с теменной корой и другими отделами мозга [39, 163]. Получает она многочисленные волокна от VI, V2 и V3 полей [193, 567, 616, 618]. В свою очередь, поле V3 обеспечивает информацией боковую внутристеночную область и принимает гномонические проекции от нее [26, 27, 31, 61, 106]. Она также связана с височными полями [70, 559, 560] и теменно-затылочной областью [115]. Обнаруживается взаимная связь и с лобным глазным полем [493, 526] и полем V4 [164].
В последнее время особое внимание нейрофизиологами обращено на поле V3A. Именно это поле обладает многочисленными связями с теменной корой и другими отделами мозга [39, 163]. Получает она многочисленные волокна от VI, V2 и V3 полей [193, 567, 616, 618]. В свою очередь, поле V3 обеспечивает информацией боковую внутристеночную область и принимает гномонические проекции от нее [26, 27, 31, 61, 106]. Она также связана с височными полями [70, 559, 560] и теменно-затылочной областью [115]. Обнаруживается взаимная связь и с лобным глазным полем [493, 526] и полем V4 [164].
Область V3A была обнаружена Zeki [568, 619, 620]. Им установлено, что многие нейроны V3A поля реагируют на отсутствие бинокулярного слияния изображения [620], изменение направления движения цели [181, 191, 618, 619]. Эти нейроны также регистрируют положение глазного яблока в глазнице [180, 181]. Многие нейроны V3A поля различают реальное движение зрительной цели от эквивалентного движения зрительного стимула в поле зрения, возникающего при движении глаза [181].
Современными исследованиями показано, что поле V3A является важным звеном между зрительными полями и сенсорно-двигательными областями теменной и лобной коры. Выявленные недавно явления активизации нейронов этого поля, упреждающие внимание животного к зрительной цели, свидетельствуют о их возможном участии в познавательных функциях [392—395].
Поле V4.Около 30 лет назад Zeki [615] выявил кортикальное поле в области лунной борозды обезьяны, которое содержало нейроны, избирательно отвечающие на стимулы различной спектральной характеристики. Это поле было названо полем V4. Сейчас оно известно как «цветовое поле» ассоциативной коры (рис. 4.2.50, 4.2.61). На это поле прямо проецируются кодирующие цвет нейроны, расположенные в тонких темных полосах V2 поля, и нейроны светлых полос. Большинство нейронов поля относятся к комплексным, ориентацион-ным и энд-стопт (end-stopped) рецептивным полям. Только небольшое их количество избирательно реагирует на направление движения объекта [144, 194, 386]. Наиболее важной их характеристикой является выраженный ответ на цветовой стимул. При повреждении поля отмечается нарушение способности распознавания объекта. Сводится это к тому, что объект видоизменяется в размере, степени перекрытия изображения и недостаточности информации о контуре изображения. Проецируется поле V4 на нижнее височное поле коры (IT) [144].
V5 (среднее височное поле).Существует, по крайней мере, два пути обработки информации экстрастриарной корой. Выявлены они путем сопоставления изменений, наблюдаемых в поведении животных после травмы, со струк-
турными изменениями коры [561]. Ungerleider и Mishkin [561] установили, что животные с повреждением височной коры хуже опознавали зрительные объекты. При этом функции зрения (острота зрения) сохранялись. Повреждение теменной коры не приводило к нарушению функции распознавания зрительного объекта, но решение пространственных задач существенно нарушалось. На основании этих данных авторы постулировали, что существует два отличающихся потока зрительной информации — височный (вентральный) поток, участвующий в опознании объекта, и дорзальный поток (теменной), участвующий в решении пространственных задач. Необходимо отметить, что эти оба тракта различны и в структурном отношении. Так, теменной поток, в первую очередь, обеспечивается магноцеллюлярными клетками, а височный — парвоцеллюлярными (см. ниже).
К одному из указанных трактов (именно к теменному, дорзальному) относится поле V5 [392]. В 1971 — 1972 гг. Allman, Kaas, а также Dubner, Zeki [152] при исследовании обезьяны выявили поля, отличающиеся своей цитоархи-тектоникой. Располагались они в среднем височном поле (МТ), отличающемся наличием большого количества миелина. Авторы предположили, что это поле является местом представительства полуполя противоположного глаза. Позже Zeki [616] обнаружил подобное поле у обезьян другого вида и назвал его V5 полем. Он отметил, что нейроны этого поля чувствительны к движущимся стимулам. Этим они напоминали комплексные клетки поля VI, но отличались большим размером рецептивных полей и разнообразием регистрируемых направлений движения цели. Некоторые нейроны реагировали на цвет.
На поле V5 (МТ, средняя височная кора) проецируются нейроны, расположенные в слое IVB поля VI и нейроны тонкой полосы V2 поля [143, 392, 507]. Благодаря своей способности реагировать на движение объекта, это поле было названо «поле движения» экстрастриарной коры.
Нейроны поля V5 (поле МТ) относятся к анализирующим движение нейронам более высокого порядка. При этом 95% нейронов поля МТ спецализированы в отношении анализа направления движения объекта [22, 183, 211, 214, 399]. Рецептивные поля этих нейронов объединяют информацию о движении объекта на большой площади. Схожие по функции нейроны содержит и первичная зрительная кора (поле VI) (см. выше), но они не способны анализировать скорость и направление движения при изменении пространственных характеристик этого движения, поскольку их рецептивные поля небольшие. Нейроны МТ поля реагируют на изменение независимо от изменения пространственных характеристик. Они, кроме того, анализируют характер движения нескольких объектов
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 при разнонаправленном их перемещении, комбинируя составляющие этого движения.
при разнонаправленном их перемещении, комбинируя составляющие этого движения.
Диапазон реакций нейронов МГ-поля довольно широкий. Анализируют они, помимо скорости и направления движения, также форму объекта, контрастность, «текстуру» и др. [559, 560].
Поле V5a (MST — среднее верхнее темпоральное поле), расположенное несколько ниже поля V5, отличается тем, что участвует в управлении движениями глаз (плавное преследование) и туловища [210, 214]. В анализе информации относительно движения объекта участвует и поле 7а (внимание и интеграция).
Второй поток (нижневисочный, вентральный) «-специализируется» на анализе формы зрительного объекта. Схематически этот поток выглядит следующим образом: VI —>• V2 --> _ V4 —<■ IT (нижнее височное поле). Поле V4 получает вход также от V3 поля и МТ (рис. 4.2.61, 4.2.62).
Клетки поля IT исключительно чувствительны к комплесным стимулам, особенно стимулам, характеризующим ориентацию, положение объекта. Обладает оно и специальными функциями, а именно, участвует в распознавании руки [183—185] и лица. Это поле обладает рецептивными полями большого размера (25 X 25°), которые получают информацию, помимо периферии сетчатки, и от макулярной области. Тем не менее в нем отсутствует ретино-топическая организация. Нарушение функций этого поля сопровождается утерей способности к распознаванию формы объекта. При этом сохраняется способность анализировать цвет и движение.
Нарушение функции ассоциативных зрительных полей и связанных с ними других участков коры приводит к развитию многообразных патологических состояний. Более подробные сведения об этих нарушениях приведены в разделе «Головной мозг».
В заключение необходимо отметить, что если физиологические закономерности реакций ассоциативной коры достаточно хорошо изучены, то механизмы зрительного восприятия как психофизиологического явления до сих пор остаются далеко не понятыми. Эти механизмы являются объектом исследования психофизиологов. Обсуждение этих вопросов выходит за рамки настоящей работы, но с ними можно ознакомиться, например, у М. Арбиб (1976) [1].
Р- и А1-тракты. Выше мы неоднократно упоминали о Я- и Af-трактах. Теперь имеет смысл более подробно их охарактеризовать.
Одним из свойств зрительного пути является его структурно-функциональная двойственность. С одной стороны, существует путь, отличающийся жесткой иерархической организацией анатомических структур. Обработка зрительной информации в этой системе характеризуется постепенным ее усложнением по мере
передачи информации от сетчатой оболочки (фоторецепция и первичная обработка) до коры головного мозга (зрительное восприятие). Эта система включает сетчатую оболочку, наружное коленчатое тело, первичную зрительную кору, ассоциативные поля (VI, V2, V3, V4 и V5) и зрительные области височной и теменной коры.
С другой стороны, в рамках этой структуры функционируют два параллельных тракта. Это так называемые Р- и М-тракты.
Первоначально необходимо отметить, что указанные два параллельных тракта в зависимости от основной функции называют «что»- и «ггдея-трактами. То есть, Я-тракт выполняет основную функцию качественного анализа изображения, а УИ-тракт—его пространственную локализацию и анализирует движение.
Начинаются оба тракта в сетчатой оболочке (ганглиозные Р- и Af-клетки сетчатки), которые проецируются на Р- и М-слои наружного коленчатого тела. Затем афферентные волокна, идущие от Р- и М-слоев наружного коленчатого тела, проецируются на IVC-слой первичной зрительной коры (рис. 4.2.50, 4.2.62, 4.2.63). Некоторые волокна проходят к слоям I и VI. При этом волокна, исходящие из парвоцеллюлярных (Р-тракт) и магноцеллюлярных слоев (Af-тракт) наружного коленчатого тела, распределяются в коре неодинаково.
Аксоны парвоцеллюлярных слоев, определяющих пространственную остроту зрения и цвет, заканчиваются в слоях IVA, IVC и VIA («капли» — цвет; нейроны, расположенные между «каплями» — форма объекта). Имеется также прямой вход от парвоцеллюлярных нейронов наружного коленчатого тела в III слой зрительной коры (рис. 4.2.63).
| 4а: |
Проекция магноцеллюлярных нейронов осуществляется на слой IVC. Клетки этого слоя анализируют информацию о пространственной ориентации и наличии движения объекта.
| Ч А 1 | х\ ■■■' ''''■■ | ||
| ! / 1 | \ | ||
| /// ^—к | |||
| ____ 1_____ Ч |
 Рис. 4.2.63. Схема проекции наружного коленчатого
Рис. 4.2.63. Схема проекции наружного коленчатого
тела на зрительную кору (по Hubel, Wiesel. 1972;
Hendriksen et al., 1978; Blasdel, Lund. 1983; Fitz-
patrick et al., 1983):
P,, Р.г, Ръ— вход от парвоцеллюлярных слоев: М — вход от магноцеллюлярных слоев. Пунктирной линией отмечены границы «глазных доминантных колонок», шириной 400—500 мкм
Функциональная анатомия зрительной системы
 IVC-слой коры взаимодействует с другими слоями, расположенными как более поверхностно, так и более глубоко. Он соединяется со II и IIIслоями, а последние слои соединяются с V слоем. В свою очередь, слой V проецируется на слои VI и IV.
IVC-слой коры взаимодействует с другими слоями, расположенными как более поверхностно, так и более глубоко. Он соединяется со II и IIIслоями, а последние слои соединяются с V слоем. В свою очередь, слой V проецируется на слои VI и IV.
После обработки в первичной зрительной коре два тракта различным путем передают информацию различным полям коры головного мозга для дальнейшего анализа. При этом пар-воцеллюлярный тракт, который можно назвать височным трактом, выглядит следующим образом: поля VI —> V2 —► V3 -* V4 —> нижнее височное поле. Этот тракт определяет возможность распознавания объекта (рис. 4.2.62).
Магноцеллюлярный тракт, в свою очередь, можно назвать теменным (париетальным) трактом, поскольку направляется он к теменной коре, но основная обработка информации происходит в средней темпоральной (МТ) и медиальной верхней темпоральной коре (MST) (рис. 4.2.62). Основной функцией этого пути является анализ характера движения зрительного объекта [141, 210].
Учитывая существование определенных функциональных особенностей Р- и Af-трактов, не является неожиданным существование различных проявлений их поражения. Изучены они в экспериментальных условиях. Различия в проявлении поражения того или иного тракта касаются двух функций — обнаружения зрительного объекта и его распознавания [561]. Именно эти функции отличают указанные два тракта. Более подробно особенности проявления повреждения того или иного тракта приведены в табл. 4.2.1.
Таблица 4.2.1. Особенности нарушения зрительных функций при повреждении парво- и магноцел-люлярных трактов
| Функция | Парвоцеллюлярный | Магноцеллю- |
| тракт | лярный тракт | |
| Цвет | Дефицит в опреде- | Отсутствие |
| лении цветов, но от- | нарушений | |
| сутствие нарушений | ||
| в обнаружении объ- | ||
| екта, основанном на | ||
| анализе различий в | ||
| длине Дата добавления: 2016-06-05; просмотров: 3861; Поиск по сайту Узнать еще

Публикации по технике и механике
Публикации по биологии
Публикации по информатике
Публикации по строительству
Публикации по физике
Публикации по химии
Публикации по электронике
Публикации по искусству
Публикации по географии
Публикации по медицине
Публикации по педагогике |