Функциональная анатомия сетчатки
В последнее время показано, что уже на уровне сетчатки происходит существенная обработка сигнала перед передачей его центральным структурам мозга [212].
Подробная структурная организация сетчатой оболочки нами приведена во второй главе. Задачей настоящего раздела является описа-
ние функционального значения нейронов сетчатки и связи между ними в процессе формирования и передаче информации другим отделам центральной нервной системы. При подобном подходе к изложению материала нам необходимо только напомнить основные ее структуры. При этом в функциональном отношении логично подразделить сетчатку на две нейронные системы — вертикальную и горизонтальную (рис. 4.2.3, см. цв. вкл.), состоящие из шести типов нейронов [592].
К вертикальной системе относятся фоторе-цепторные клетки (палочки и колбочки), биполярная клетка и ганглиозная клетка. Именно в этой цепи нервный импульс, сформированный в фоторецепторе под действием света, передается головному мозгу посредством зрительного нерва.
Горизонтальная система нейронных связей состоит из нейронов, определяющих связь между фоторецепторами, биполярными и ганглиоз-ными клетками в плоскости сетчатки. К этим нейронам относятся горизонтальные, амакрино-вые и межплексиформные клетки. Именно го-
Функциональная анатомия зрительной системы
 ризонтальные связи объясняют многие нейрофизиологические закономерности функционирования сетчатки.
ризонтальные связи объясняют многие нейрофизиологические закономерности функционирования сетчатки.
Перед тем как остановиться на описании нейронных связей сетчатки необходимо дать хотя бы краткую характеристику основным физиологическим понятиям функционирования зрительной системы, без которых дальнейшее восприятие материала будет затруднительно.
Многие закономерности анализа изображения были установлены психологами задолго до выявления нейрофизиологами и нейрогисто-логами особенностей формирования нейронных сетей сетчатки и структур более высоких уровней организации зрительного анализатора. Психологами показано, что в процессе восприятия и распознавания изображения наибольшее значение имеет выделение и анализ контуров объекта, наличие и характер перекрещивающихся линий и углов, фоновое окружение объекта, наличие признаков его движения, а также спектральные характеристики отраженного от объекта света. Возможность регистрации и анализа перечисленных признаков предполагает наличие в сетчатой оболочке сложных детекторов приведенных характеристик. Причем изучены и основные закономерности функционирования подобных детекторов. Один из подобных детекторов определяет существование так называемого «пространственного контраста» (пространственная оппонентность). Наиболее убедительно существование явления «пространственного контраста» проявляется на тестах типа «решетки Геринга», полос Маха и явления восприятия объекта более светлым на темном фоне и наоборот (рис. 4.2.4). Рассматривая две окружности, расположенные на чер-
ном и белом фоне, возникает впечатление того, что круг на белом фоне более темный, чем круг на черном фоне. В то же время интенсивность окрашивания этих кругов одинаковая. Близкая закономерность выявляется при рассматривании полос Маха. При длительном рассматривании этого рисунка возникает впечатление, что по диагонали белых полос между черными квадратами располагаются серые фигуры четырехугольной формы.
Приведенные примеры, во-первых, указывают на то, что при восприятии изображения большое значение имеет фон, окружающий анализируемый объект. Во-вторых, становится понятным, что детекторы, воспринимающие изображение, особенно границу между двумя участками различной плотности окрашивания, находятся в оппонетных отношениях. Физиологическое значение этого явления сводится к существованию механизмов усиливания контраста именно на границе перепада интенсивности окрашивания, что способствует выделению контуров объекта. Именно это явление легло в основу физиологического понятия «латеральное торможение», которое реализуется благодаря наличию обратных связей между нейронами, обладающими тормозным свойством и организованными в «рецептивные поля» (см. ниже). Таким образом, существование латерального торможения связано с тем, что активность каждого отдельного нейрона в цепи зависит от активности соседних клеток. Причем они находятся в оппонентных отношениях.
Психологами также выявлено существование так называемого эффекта сенсорного последствия, в нашем случае последействия зрительного сигнала. В работе зрительного анали-


Рис. 4.2.4. Психофизиологические тесты, указывающие на наличие в работе зрительного анализатора явлений:
а — оппоненции (полосы Маха); б — пространственного контраста (решетка Геринга); в — значения фона при восприятии изображения
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 затора этот эффект проявляется в виде так называемого «последовательного образа». Наиболее ярким примером существования такого эффекта может служить следующее явление. Если после длительного рассмотрения красного изображения перевести взляд на белую поверхность, белая поверхность будет казаться зеленой, т. е. цвета, дополнительного к красному. Особенно часто приходится сталкиваться с явлением последействия движения. Например, при вращении колеса со спицами с определенной скоростью возникает впечатление вращения колеса в обратном направлении. Явление последействия указывает на существование одновременно двух антагонистических сенсорных систем, уравновешенных в обычных условиях. Можно предположить, что при длительном возбуждении одной из систем световым сигналом ее чувствительность снижается, а антогонисти-ческая система сохраняет свою чувствительность. Если предположить, что детектор красного цвета в приведенном выше примере спарен с детектором зеленого цвета, то оба они одинаково реагируют на белый цвет и на выходе пары нет ничего, кроме фона. Предположим теперь, что глаз фиксирует красную поверхность в течение некоторого времени, в результате чего рецепторы красного цвета «утомляются». Если перевести взгляд на белую поверхность, то рецепторы зеленого будут реагировать нормально, а рецепторы красного дадут ослабленную реакцию. Таким образом, свет будет восприниматься как зеленый. Необходимо отметить, что приведенное объяснение легло в основу оппонентной теории восприятия цвета, на чем мы остановимся несколько ниже. Наличие детекторов, представляющих собой организованные определенным образом рецептивные поля и обладающих антагонистическими свойствами, которые реагируют на стимулы различной формы (линии, щели, углы и т. п.) и цвет, было позднее обнаружено нейрофизиологами на уровне сетчатки, наружных коленчатых тел и различных участков коры головного мозга при помощи разработанных методов электрофизиологии, позволяющих снимать потенциал отдельного нейрона. На основе этих исследований теоретически разработаны схемы нейронных цепей, удовлетворяющие требованиям детекторов различных характеристик изображения при его анализе. В последующем ней-рогистологи при помощи световой, электронной микроскопии и методов гистохимии выявили особенности этих цепей, заключающиеся в определенной связи между нейронами различных типов, характере контактов между ними, а также в использовании нейронами при передаче нервного импульса различных нейромедиато-ров. Основополагающим понятием, возникшим в результате приведенных выше наблюдений, явилось понятие «рецептивное поле». Ранее мы неоднократно упоминали термин «рецептивное
затора этот эффект проявляется в виде так называемого «последовательного образа». Наиболее ярким примером существования такого эффекта может служить следующее явление. Если после длительного рассмотрения красного изображения перевести взляд на белую поверхность, белая поверхность будет казаться зеленой, т. е. цвета, дополнительного к красному. Особенно часто приходится сталкиваться с явлением последействия движения. Например, при вращении колеса со спицами с определенной скоростью возникает впечатление вращения колеса в обратном направлении. Явление последействия указывает на существование одновременно двух антагонистических сенсорных систем, уравновешенных в обычных условиях. Можно предположить, что при длительном возбуждении одной из систем световым сигналом ее чувствительность снижается, а антогонисти-ческая система сохраняет свою чувствительность. Если предположить, что детектор красного цвета в приведенном выше примере спарен с детектором зеленого цвета, то оба они одинаково реагируют на белый цвет и на выходе пары нет ничего, кроме фона. Предположим теперь, что глаз фиксирует красную поверхность в течение некоторого времени, в результате чего рецепторы красного цвета «утомляются». Если перевести взгляд на белую поверхность, то рецепторы зеленого будут реагировать нормально, а рецепторы красного дадут ослабленную реакцию. Таким образом, свет будет восприниматься как зеленый. Необходимо отметить, что приведенное объяснение легло в основу оппонентной теории восприятия цвета, на чем мы остановимся несколько ниже. Наличие детекторов, представляющих собой организованные определенным образом рецептивные поля и обладающих антагонистическими свойствами, которые реагируют на стимулы различной формы (линии, щели, углы и т. п.) и цвет, было позднее обнаружено нейрофизиологами на уровне сетчатки, наружных коленчатых тел и различных участков коры головного мозга при помощи разработанных методов электрофизиологии, позволяющих снимать потенциал отдельного нейрона. На основе этих исследований теоретически разработаны схемы нейронных цепей, удовлетворяющие требованиям детекторов различных характеристик изображения при его анализе. В последующем ней-рогистологи при помощи световой, электронной микроскопии и методов гистохимии выявили особенности этих цепей, заключающиеся в определенной связи между нейронами различных типов, характере контактов между ними, а также в использовании нейронами при передаче нервного импульса различных нейромедиато-ров. Основополагающим понятием, возникшим в результате приведенных выше наблюдений, явилось понятие «рецептивное поле». Ранее мы неоднократно упоминали термин «рецептивное
поле». Об особенностях функционирования рецептивных полей мы сейчас остановимся более подробно.
Рецептивное полена уровне сетчатки представляет собой совокупность точек сетчатки (фоторецепторных клеток), в пределах которой зрительный стимул вызывает возбуждение или торможение одного нейрона следующего (более высокого) порядка, в частности ганглиозной клетки.
Еще в 30-е годы прошлого столетия Hart-line [231, 232] развил концепцию рецептивного поля при изучении глаза краба и лягушки. В дальнейшем были выполнены классические эксперименты на кошках [319, 320], сформировавшие базу для последующих исследований зрительной системы млекопитающих.
Как было указано выше, возможность подобных экспериментов появилась в связи с созданием методик подведения микроэлектрода непосредственно к одному нейрону. Именно таким образом регистрируется потенциал действия ганглиозных клеток сетчатки, нейронов наружных коленчатых тел и нейронов зрительной коры головного мозга во время освещения сетчатки паттернами различной формы и плотности энергии (рис. 4.2.5). Это позволило исследователям анализировать как параметры зрительных стимулов, возбуждающих или тормозящих нейроны, так и особенности реакции нейронной системы.
Используя эту методику, первое, что обнаружили Kuffler и Hartline [320], это существование двух категорий ганглиозных клеток. Клетки ON-типа возбуждались при освещении, а OFF-типа возбуждались при отсутствии освещения. Причем участки сетчатки, отвечающие противоположной реакцией, располагались рядом, чаще в виде кольца. Центральная часть такой окружности отвечала на стимул одним типом реакции, а периферия противоположной. При регистрации потенциала действия ганглиозной клетки световое пятно, помещенное
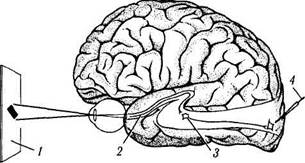
Рис. 4.2.5. Схема проведения электрофизиологических исследований функциональной активности различных отделов зрительного пути (объяснения в тексте):
1 — зрительное поле; 2 — зрительный тракт; 3 — наружное коленчатое тело; 4 — электрод в зрительной коре
Функциональная анатомия зрительной системы

 в центр рецептивного поля, вызывает усиление активности клетки, т. е. формируется ON-ответ (рис. 4.2.6, см. цв. вкл.). Когда такое же небольшое световое пятно помещают вне рецептивного поля, нейрон уменьшат свою активность (OFF-ответ). Подобного типа рецептивное поле называется ON-центр рецептивным полем. Существуют и OFF-центр рецептивные поля, особенностью которых является торможение ганглиозной клетки при освещении центра рецептивного поля.
в центр рецептивного поля, вызывает усиление активности клетки, т. е. формируется ON-ответ (рис. 4.2.6, см. цв. вкл.). Когда такое же небольшое световое пятно помещают вне рецептивного поля, нейрон уменьшат свою активность (OFF-ответ). Подобного типа рецептивное поле называется ON-центр рецептивным полем. Существуют и OFF-центр рецептивные поля, особенностью которых является торможение ганглиозной клетки при освещении центра рецептивного поля.
Куфлером изучались также рецептивные поля с целью определения «оптимального сигнала», т. е. сигнала в наибольшей степени возбуждающего ганглиозную клетку. Оказалось, что самым сильным сигналом для ON-центр ганглиозной клетки было световое пятно, полностью выполняющее центр рецептивного поля (см. рис. 4.2.6). Таким же образом наиболее оптимальным для тормозного сигнала явилось освещение периферии рецептивного поля стимулом, имеющим вид кольца. После подобного торможения в момент снятия светового сигнала нейрон формировал потенциал действия.
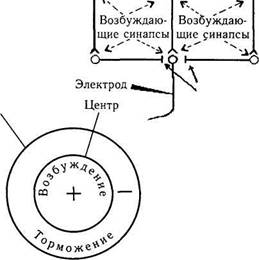
Установлено также, что если освещать рецептивное поле большим световым пятном, накрывающим как центр, так и периферию поля, возникает значительно более слабый ответ, чем при использовании маленького пятна, освещающего только центр. Таким образом, тормозная реакция периферии рецептивного поля ослабляла или даже устраняла центральное возбуждение. Приведенные выше реакции рецептивного поля связаны с особым типом обратных связей тормозного характера между нейронами сетчатой оболочки (рис. 4.2.7).
Из приведенных выше фактов становится ясно, что в основе большинства электрофизиологических особенностей рецептивных полей лежат явления возбуждения и торможения, морфологическим субстратом которых являются обратные связи возбуждающего или тормозного характера между рядом расположенными нейронами. Именно наличием таких связей между нейронами с формированием рецептивных полей можно объяснить феномены, приведенные на рис. 4.2.4.
В настоящее время известно большое количество разнообразных типов реакций зрительной системы, основой которых является функционирование рецептивных полей. Существование рецептивных полей сетчатки определяет фундаментальные функциональные ее свойства, такие как обеспечение одновременного и последовательного зрительного контраста, острота зрения, движение объекта, световая и темно-вая адаптация, обработка информации о цвете объекта. При этом указывают на наличие довольно сложной организации рецептивных полей, выполняющих ту или иную функцию. На уровне сетчатки доказано наличие рецептивных полей, реагирующих на форму объекта (выде-
|
| Тормозные синапсы |
| Окружение |
ии
Рис. 4.2.7. Схема обратных свезей между нейронами сетчатки, формирующими ргцептивные поля (объяснение в тексте)
ляя такие признаки, как наличие и длина линий, наличие углов и т. п.), его движение, спектральную характеристику объекта. Благодаря сложному взаимодействию нейронов уже в сетчатке зрительная информация обрабатывается, кодируется и по зрительному нерву передается наружному коленчатому телу [592]. Структура рецептивных полей сохраняется и на уровне наружных коленчатых тел, а также в зрительной коре головного мозга. Именно подобная организация обеспечивает передачу информации по принципу «точка к точке». Этот принцип сводится к тому, что фоторецепторы определенных участков сетчатой оболочки передают информацию ганглиозным клеткам, а затем нейронам наружного коленчатого тела и зрительной коры, сохраняя при этом топографическое пространственное расположение рецептивных полей. Это топографическое распределение не линейное. Примером тому является значительно большее представительство макулярной области в наружном коленчатом теле и коре относительно остальной части сетчатки.
В настоящее время достаточно точно выявлены морфологические основы нескольких нейронных сетей сетчатки, определяющие их различные функции. Это система палочек, система колбочек, «карликовая» система фовео-лярной области, тракт S-колбочек и обратные связи. Особенности организации этих сетей сводятся к наличию определенных типов нейронов, а также синаптических соединений, использующих различные медиаторы.
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 Теперь мы кратко охарактеризуем каждую из указанных систем.
Теперь мы кратко охарактеризуем каждую из указанных систем.
Нейронная система палочек.Палочки и связанные с ними нейроны сетчатки ответственны за функционирование трактов, обеспечивающих «ночное зрение». То есть эта система нейронов, повышая чувствительность зрительной системы, позволяет анализировать зрительные объекты при низком освещении (скотопи-ческие условия).
Несмотря на то, что у человека существует «центральное зрение», определяющее высокую разрешающую способность зрительного анализатора, и цветовое зрение, система палочек также важна. При патологии нейронов этой системы развивается ночная слепота, наиболее часто встречающаяся при пигментном ретините.
Показано, что в сетчатке человека преобладают палочки, за исключением фовеолы. Их концентрация достигает максимума в кольце вокруг фовеа, шириной приблизительно 5 мм [405]. Высокая плотность фоторецепторов обеспечивает, в значительной степени, остроту зрения (см. главу 3). Максимум поглощения световой энергии пигментом палочки располагается в области 497 нм.
Каким же образом организована система палочек?
Сигнал, сформированный в момент освещения, от палочки передается биполярным клеткам. Выявляется только один морфологический тип биполярной клетки, который формирует си-наптическую связь с палочкой (синапс ленточ-
ного типа). Эта биполярная клетка одномоментно получает информацию от 15—30 палочек, что является морфологической основой конвергенции сигнала. Происходит это в наружном плексиформном слое (рис. 4.2.3; 4.2.8, см. цв. вкл.). Затем биполярные клетки палочек посылают аксоны в направлении внутреннего плексиформного слоя, где они заканчиваются в глубоких его слоях вблизи тел ганглиозных клеток. При этом прямого контакта между биполярной и ганглиозной клетками нет, а взаимодействие между ними осуществляется посредством амакриновых клеток. Вся обработка информации происходит на уровне дендритов амакриновых клеток (внутренний плексиформ-ный слой).
Функции большинства типов амакриновых клеток изучены пока недостаточно. Тем не менее известно, что они обеспечивают антагонистическое окружение рецептивного поля, благодаря механизму латерального торможения. Подобная организация нейронов обеспечивает дивергенцию и конвергенцию сигнала палочек на уровне биполярных клеток палочек и амакриновых клеток перед тем, как информация поступает ганглиозной клетке.
Формирование рецептивного поля осуществляется несколькими типами амакриновых клеток (All, A17).
Амакриновая клетка АН типа характеризуется тем, что при помощи щелевого соединения (электрический синапс) она соединяется с биполярной клеткой колбочки (рис. 4.2.8, 4.2.9).
Палочки
Колбочки
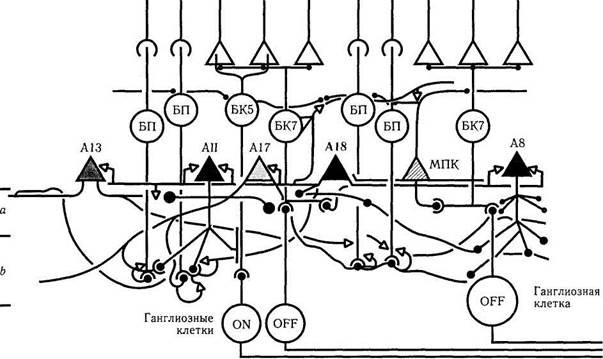
Рис. 4.2.9. Особенности связей амакриновой клетки типа А18 (объяснение в тексте)
Функциональная анатомия зрительной системы
 Последняя, в свою очередь, вступает в контакт с ганглиозной клеткой колбочки, что происходит в субслое b внутреннего плексиформного слоя [314]. В результате этого формируется так называемый «возвратный синапс», известный как реципроктный синапс. Эти ганглиоз-ные клетки отвечают на освещение деполяризацией, формируя ON-центр рецептивного поля. В то же время амакриновые клетки АН, контактирующие с ганглиозной клеткой при помощи синаптической ленты в субслое а, отвечают на освещение сетчатки гиперполяризацией, образуя OFF-центр рецептивного поля (рис. 4.2.8, 4.2.9). Именно благодаря этому образуются рецептивные поля ON/OFF-типа.
Последняя, в свою очередь, вступает в контакт с ганглиозной клеткой колбочки, что происходит в субслое b внутреннего плексиформного слоя [314]. В результате этого формируется так называемый «возвратный синапс», известный как реципроктный синапс. Эти ганглиоз-ные клетки отвечают на освещение деполяризацией, формируя ON-центр рецептивного поля. В то же время амакриновые клетки АН, контактирующие с ганглиозной клеткой при помощи синаптической ленты в субслое а, отвечают на освещение сетчатки гиперполяризацией, образуя OFF-центр рецептивного поля (рис. 4.2.8, 4.2.9). Именно благодаря этому образуются рецептивные поля ON/OFF-типа.
Необходимо отметить, что амакриновые клетки АИ также соединяются между собой. Это происходит в субпластинке b при помощи щелевых контактов. Кроме того, амакриновая клетка АН соединяется с биполярной клеткой колбочки. Благодаря приведенной выше схеме нейронных связей амакриновой клетки АН с биполярной клеткой колбочки и ганглиозными клетками сигналы, исходящие от палочек, используются и колбочковой системой при изменении степени освещенности (темновые и мезо-пические условия).
Вторым типом амакриновых клеток являются клетки типа А17. Эти клетки относятся к ГАМК-эргическим нейронам [425, 592] и обладают широким дендритным полем, что позволяет одной клетке объединить до 1000биполярных клеток палочек. Дендриты амакриновых клеток А17 достигают субпластинки b внутреннего плексиформного слоя, где и завершаются (рис. 4.2.8, 4.2.9). Эти клетки не образуют синапсов с другими амакриновыми и ганглиозными клетками и их функцией является только объединение биполярных клеток палочек, обеспечивая конвергенцию сигнала. По этой причине амакриновая клетка А17 обладает самостоятельным рецептивным полем и регулирует уровень чувствительности палочек и биполярных клеток по площади сетчатки, участвуя тем самым в темновой и световой адаптации. Благодаря конвергенции сигнала эта клетка очень чувствительна к освещению низкой интенсивности.
Третьим типом амакриновых клеток тракта палочек является клетка типа А18, дендритное поле которой распределяется только в субпластинке а внутреннего плексиформного слоя (рис. 4.2.9). Эта амакриновая клетка относится к допаминэргическим (выявляется при проведении иммуногистохимических реакций на ферменты синтеза допамина, тирозин-гидроксила-зы). Отростки клетки типа А18 окружают тела и дендриты амакриновых клеток типа АИ, А8, А17 (рис. 4.2.9). Они также образуют большое количество синапсов с трактами палочек, амакриновыми и биполярными клетками колбочек. Эти амакриновые клетки отвечают на возбуж-
дение деполяризацией. Предполагают, что до-паминэргическая клетка А18 увеличивает размер рецептивного поля, повышая при этом чувствительность ганглиозных клеток в ското-пических условиях. Увеличение рецептивного поля исключительно важно (помимо изменения скорости синтеза пигментов фоторецепторов) в реализации механизмов темновой адаптации.
Система приведенных выше амакриновых клеток обеспечивает не только конвергенцию сигнала, но и его дивергенцию, благодаря обратным связям. Степень конвергенции и дивергенции системы палочек можно проиллюстрировать на примере организации сетчатки кошки [314]. Конвергенция сигнала сводится к тому, что приблизительно 1500 палочек передают информацию отдельной ON-бета ганглиозной клетке посредством 100 биполярных клеток палочек, 5 амакриновых клеток типа АИ и 4 биполярных клеток колбочек. Информация к OFF-альфа ганглиозной клетке поступает от 75 000 палочек, 5000 биполярярных клеток палочек и 250 амакриновых клеток. Дивергенция палочковой системы сводится к тому, что отдельная палочка передает информацию двум биполярным клеткам, а от них 5 амакриновым клеткам типа АИ, 8 биполярным клеткам колбочек и двум ON-бета ганглиозным клеткам. Благодаря существованию дивергентной и конвергентной систем обеспечивается объединение и усиление сигнала палочек при очень низком освещении. Чувствительность столь высока, что зрительная система реагирует на единственный квант световой энергии. Благодаря наличию связи между палочковой и колбочковой системами посредством амакриновых клеток, палочковая нейронная система в мезопи-ческих условиях передает свои функции кол-бочковой системе.
Необходимо подчеркнуть и то, что приведенная выше схема взаимодействия между нейронами палочкового тракта является основой формирования рецептивных полей, обеспечивающих наиболее важные функции зрительной системы — контрастную чувствительность, адаптацию.
Нейронная система колбочек.Нейронная система колбочек обеспечивает наибольшую остроту зрения в фотопических условиях, а также обладает способностью цветовосприятия. В определенных условиях (темновые) колбоч-ковая система регистрирует ахроматические сигналы. Первоначально мы охарактеризуем основные закономерности формирования кол-бочковой системы.
Необходимо отметить, что система прохождения сигналов от колбочки к ганглиозным клеткам довольно существенно отличается от нейронной системы палочек. Так, одна колбочка образует синапсы с двумя биполярными клетками, в то время как палочка только с одной (рис. 4.2.3, 4.2.8). Благодаря этому уже
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 в наружном плексиформном слое формируются различные тракты колбочковой системы [363, 592]. Кроме того, биполярные клетки колбочек контактируют непосредственно с ганглиозны-ми клетками. В результате такой структурной организации тракты колбочек сетчатки человека существуют в виде двух параллельных каналов передачи информации, идущей непосредственно от колбочки к ганглиозной клетке, используя прямой путь — биполярную клетку.
в наружном плексиформном слое формируются различные тракты колбочковой системы [363, 592]. Кроме того, биполярные клетки колбочек контактируют непосредственно с ганглиозны-ми клетками. В результате такой структурной организации тракты колбочек сетчатки человека существуют в виде двух параллельных каналов передачи информации, идущей непосредственно от колбочки к ганглиозной клетке, используя прямой путь — биполярную клетку.
Какова целесообразность существования таких двух параллельных каналов? Подобная организация позволяет одному каналу обеспечивать информацией ганглиозную клетку о большей яркости в центре (ON-центр рецептивного поля), а второй, наоборот, о большей яркости по периферии (OFF-центр рецептивного поля). Именно благодаря этому свойству и формируется нейрофизиологическая основа одновременного контраста.
Анатомическим субстратом ON- и OFF-ка-налов является тип синапса между колбочкой и биполярной клеткой. Гиперполяризующий тип биполярной клетки формирует OFF-центр канал, а деполяризующий — ON-центр канал (рис. 4.2.8). Таким образом, тип сигнала, поступающего ганглиозной клетке (ON- или OFF-центр), определен типом биполярной клетки [363, 592].
Каким образом функционируют эти каналы? Колбочки при освещении возбуждаются. Причем одна биполярная клетка деполяризуется, а другая гиперполяризуется. В результате этого «расщепляется» первоначальный сигнал колбочки на «светлый» (ON-центр) или «темный» (OFF-центр). Эти реакции биполярных клеток передаются непосредственно ганглиозным клеткам. Один канал идет к гаглиозным клеткам, дендриты которых располагаются в субпластинке Ь внутреннего плексиформного слоя и становятся ON-центром, а другой — к дендритам ганглиозных клеток в субпластинке а, становящимся OFF-центром. Одновременный контраст обеспечивается взаимодействием двух этих каналов благодаря механизму латерального торможения. Структурно латеральное торможение обеспечивается горизонтальными клетками (рис. 4.2.3, 4.2.8). При этом отрицательный синапс обратной связи между горизонтальной клеткой и колбочкой в специализированном синапсе («триада)» позволяет большое рецептивное поле горизонтальных клеток (горизонтальные клетки соединены электрическими синапсами) сузить и окружить зоной обратной реакции. Подобная концентрическая организация затем передается биполярным клеткам, а от них ганглиозной клетке.
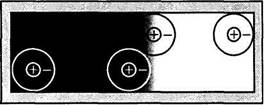
Исходя из функциональной организации рецептивных полей ганглиозных клеток, можно представить следующее объяснение явления одновременного контраста (рис. 4.2.10). Величина возбуждения нейронов сетчатки на стимул

|
Свет
h
о яО
Темно
Расстояние от границы
Рис. 4.2.10. Электрофизиологическая реакция рецептивного поля на границе различной степени освещенности
в виде границы темное/светлое зависит от положения этой границы в их рецептивных полях. Ответ нейрона максимален, когда граница темное/светлое пересекает рецептивное поле в точке между центром рецептивного поля и его периферией. Поэтому максимальное возбуждение OFF-нейронов наблюдается в том случае, если центры их рецептивных полей расположены около границы, но с темной ее стороны. Напротив, максимальное возбуждение нейронов с ON-центром наблюдается в том случае, когда их центры рецептивных полей располагаются со светлой стороны границы. Если отмечать среднюю величину возбуждения всех нейронов с ON- и OFF-центрами в зоне границы темное/светлое, то полученное пространственное распределение будет соответствовать кривой воспринимаемого одновременного контраста.
Механизм, лежащий в основе явления одновременного контраста, обеспечивает частичную функциональную компенсацию погрешностей диоптрического аппарата глаза и является важным для восприятия формы объекта.
Тракт «карликовых» клеток и цветовое зрение.Колбочковый («карликовый») тракт, начинающийся в области центральной ямки, обеспечивает максимальную разрешающую способность зрительной системы, а также цветовое зрение.
Перед тем как описать нейронные сети, обеспечивающие цветовое зрение, имеет смысл кратко остановиться на современных представлениях относительно физиологии цветового зрения. Лишь затем мы опишем структурное обеспечение цветовых функций.
Цвет является одним из свойств объектов окружающего нас мира, воспринимаемое как осознанное зрительное ощущение. Таким образом, цвет является чисто сенсорным понятием. У человека понятие цвета возникает в резуль-
Функциональная анатомия зрительной системы

 тате зрительного опыта. При этом возникающему ощущению придаются определенные названия, известные нам как названия цветов и их оттенков. Возникающие цветовые ощущения независимы от этнических или культурных особенностей популяции людей и обладают замечательным постоянством. Выражается это лингвистическим сходством при обозначении того или иного цвета у разных народов. Тем не менее необходимо указать и на то, что восприятие цвета у довольно большой части здоровых людей (85%) может довольно существенно отличаться. Это может быть связано как с особенностями «цветовой культуры», иногда довольно существенно отличающейся у разных народов, так и с наличием полиморфизма ДНК, кодирующей последовательность апопротеинов колбочковых фотопигментов. Особенно существенны различия цветовосприятия при оценке различий между красными и зелеными оттенками светового спектра.
тате зрительного опыта. При этом возникающему ощущению придаются определенные названия, известные нам как названия цветов и их оттенков. Возникающие цветовые ощущения независимы от этнических или культурных особенностей популяции людей и обладают замечательным постоянством. Выражается это лингвистическим сходством при обозначении того или иного цвета у разных народов. Тем не менее необходимо указать и на то, что восприятие цвета у довольно большой части здоровых людей (85%) может довольно существенно отличаться. Это может быть связано как с особенностями «цветовой культуры», иногда довольно существенно отличающейся у разных народов, так и с наличием полиморфизма ДНК, кодирующей последовательность апопротеинов колбочковых фотопигментов. Особенно существенны различия цветовосприятия при оценке различий между красными и зелеными оттенками светового спектра.
Восприятие цвета зависит от множества параметров, включая спектральный состав света, отраженного от объекта, цветового фона, состояния световой адаптации и др.
Воспринимаемый человеком свет и, естественно, цветовая гамма в физическом смысле представляет собой исключительно небольшую часть электромагнитного излучения (рис. 4.2.11, а, см. цв. вкл.). Возникает вопрос — каковы механизмы цветового восприятия на уровне сетчатки и вышележащих отделов зрительного анализатора? Этот вопрос волновал ученых на протяжении многих столетий. Лишь после наблюдения Исаака Ньютона, установившего, что белый солнечный свет, проходя через призму, расщепляется на спектр цветов, появилась возможность создавать гипотезы механизмов цветовосприятия. Именно ему принадлежала первая теория цветового зрения. Он предположил, что восприятие цвета человеком связано с отдельными компонентами спектра, попадающего в глаз. Первоначально существовало мнение о том, что в глазу есть бесконечное множество специальных рецепторов, воспринимающих весь спектр полихромно-го цвета. Исследование цветового восприятия показывает, что эта теория не отражает действительности. Так, в спектре цветов, получаемых с помощью призмы, отсутствуют некоторые воспринимаемые нами цвета (например, коричневый, розовый, пурпурный и др.). Установлено также, что при смешении двух монохроматических пучков мы видим не два разных цвета, а один новый цвет — результат смешения. Более того, при смешении некоторых монохроматических пучков мы видим белый цвет. Подобные монохроматические цвета называют дополнительными. Именно существование дополнительных цветов дает подход к рациональному описанию механизмов цветового зрения человека.
| Зеленый | ||
| Зеленовак | з- 500 | Желтовато- |
| синий | 494 .,---- 1—^. | зеленый |
| "*•< 570 | ||
| / | л^-572 Желтый | |
| 490 V | \575 | |
| 489 / | \ | |
| Синевато- 480 -j- | 9 | т 580 Красновато |
| зеленый 1 | Белый | / желтый |
| Фиолетовый \ | Т590 | |
| 470 \ | 400— 650— / 600 Желтовато- | |
| 460 ■ | ,<440 7С | '^oo fiin красный |
| "~----------- -'' 640 | ||
| Пурпурный | Красный |
Рис. 4.2.12. Цветовая диаграмма. По окружности указаны названия цветов и соответствующие им длины волн (объяснение в тексте)
Создано большое количество систем, описывающих физические характеристики дополнительных цветов и результата их смешения (цветовое тело). Наиболее часто используют треугольник или круговую диаграмму (рис. 4.2.12). На круговой диаграмме дополнительные пары цветов можно найти, соединив любые две диаметрально противоположные точки окружности. Чтобы получить из двух цветов третий, необходимо найти на круге две соответствующие этим цветам точки и соединить их прямой линией. Любая смесь двух таких цветов дает третий цвет, который будет располагаться на этой линии. Точное положение третьего цвета зависит от интенсивности каждого из двух цветов в смеси. Аналогично можно предположить о цвете, возникающем при смешании трех монохроматических цветов, при этом он будет располагаться внутри треугольника, образованного линиями, соединяющими монохроматические цвета.
Концепция цветового круга вобрала в себя многие феномены цветового зрения. Но эта концепция позволяет лишь описать явления, не объясняя их. Она показывает только, как при смешении первичных цветов может получиться любой воспринимаемый нами цвет. Правила цветового круга приложимы к смесям света различной окраски (длины волны), но отличаются от правил при смешивании красок. Для определения вероятного цвета при смешении двух красок надо определить, какие длины волн будут поглощаться данной смесью. Поскольку краска образует цвет, отнимая часть спектра, смеси красок называют субтрактив-ными. Смешение же цветов с помощью света называют аддитивным.
Закономерности смешения цветов легли в основу трехкомпонентной теории зрения. Впервые трехкомпонентную теорию цветовосприятия предложил Томас Янг в 1801 г. Он считал, что поскольку в сетчатке не может быть бесконечного числа рецепторов, чувствительных
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
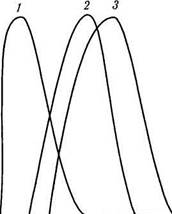
к бесконечному числу возможных вариантов оттенков цвета, наиболее логично предположить существование трех основных рецепторов, воспринимающих красный, синий и зеленый цвета. Последующие исследования Гельмгольца и Максвелла показали, что психофизиологические реакции человека на цветовые стимулы наиболее полно можно описать существованием рецепторов, реагирующих на три различные длины волны. Лишь в XX столетии благодаря разработке метода микроспектрофотометрии были выделены три типа колбочек, каждый из которых содержал особый пигмент, обусловливающий избирательность по отношению к цвету. Различные пигменты по-разному поглощают свет, обладая избирательностью по отношению к определенным длинам волн (рис. 4.2.13). Один класс зрительных пигментов имеет спектральную чувствительность, которая достигает максимума от 445 до 450 нм, другой — от 525 до 535 нм; максимум поглощения третьего пигмента лежит в области 555—570 нм. Трем пигментам соответствуют три типа рецепторов, обусловливающих нормальное цветовое зрение человека. Из описания чувствительности этих первичных рецепторов можно вывести почти все феномены цветового зрения человека. Например, из концепции круга следует, что смесь зеленого (520 нм) и красного (620 нм) должна быть неотличима от желтого с длиной волны 564 нм.
Необходимо отметить, что, используя цветовой круг, возможно описать и цвета последовательных образов. Напомним, что последовательный образ возникает в тех случаях, когда наблюдатель длительно смотрит на поверхность одного цвета, а затем переводит взгляд на белую поверхность. При этом возникает восприятие цвета, соответствующего дополнительному цвету, на который ранее смотрел наблюдатель. Цвет последовательного образа в цве-
|
§1
| 5о |
!
S
я о
§•
| 300 400 500 600 Длина волны, нм |
Рис. 4.2.13. Кривые поглощения световой энергии «синими» (/), «зелеными» (2) и «красными» (3) колбочками
товом круге лежит напротив цвета, который первоначально наблюдался наблюдателем. Последовательные образы подчиняются правилу взаимности. Если синий цвет дает последовательный образ желтого цвета, то желтый дает последовательный образ синего цвета. Приведенные выше наблюдения явились основой создания теории оппонентных процессовв цветовом зрении, которую предложил еще в 1878 г. Эвальд Геринг, а развил Харинг.
В соответствии с этой теорией цвета смешиваются по парам дополнительных цветов: синий с желтым, красный с зеленым и белый с черным (рис. 4.2.11, б). Оценка яркости выводится на основе реакции механизма «ч
Дата добавления: 2016-06-05; просмотров: 2819;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
