Базальные ганглии. Морфофункциональная организация. Функции
Базальные ганглии являются структурами ядерного типа. Они расположены внутри больших полушарий между лобными долями и промежуточным мозгом. Базальные ганглии относятся к собственно подкорковым образованиям мозга в самом узком смысле этого понятия и включают в себя три парных образования: неостриатум, паллидум (бледный шар) и ограда (claustrum).Неостриатум состоит из двух ядер: хвостатого и скорлупы (n. caudatus, putamen). Неостриатум является филогенетически новой структурой. Наиболее отчетливо он представлен начиная с рептилий. Скорлупа и хвостатое ядро по происхождению, нейронному строению, ходу проводящих путей и нейрохимическому составу являются сходными. Оба ядра, по существу, представляют собой два тяжа серого вещества, разделенных почти на всем протяжении волокнами внутренней капсулы. Паллидум, бледный шар (globus pallidum), в отличие от неостриатума, является филогенетически более древним образованием; его гомолог обнаруживается уже у рыб. Ограда расположена между скорлупой и островковой корой. Филогенетически ограда является самым новым образованием. У ежей и некоторых грызунов ее еще нет.
Морфофункциональные связи базальных ганглиев. Неостриатум образует связи с бледным шаром. Аксоны клеток неостриатума очень тонкие, до 1мкм, поэтому проведение возбуждения от неостриатума к паллидуму медленное. Стриапаллидарные волокна образуют в основном аксо-дендритные синапсы. Неостриатум оказывает двойственное влияние на нейроны паллидума – возбуждающее и тормозное. Неостриатум посылает прямые эфференты не только к паллидуму, но и к черной субстанции. Стрионигральные связи по своей природе моносинаптические и двусторонние. Большой интерес представляет обратная связь – от черной субстанции к неостриатуму. Считается, что аксоны нейронов черной субстанции, которые конвергируют к нейронам хвостатого ядра и к скорлупе, обеспечивают транспорт дофамина, синтезирующийся в нейронах черной субстанции. В неостриатуме он концентрируется в расширенных аксонных терминалях. Скорость транспорта дофамина по аксонам от черной субстанции к хвостатому ядру составляет приблизительно 0,8 мм в 1 час. Содержание дофамина в неостриатуме чрезвычайно велико. Имеются указания на то, что дофамина в неостриатуме млекопитающих в 6 раз больше, чем в паллидуме и передней части больших полушарий, в 19 раз больше, чем в мозжечке. Предполагается медиаторная роль этого амина в данной структуре. Кроме того, высказывается мнение о том, что дофамин активирует тормозные интернейроны неостриатума и таким образом подавляет деятельность его клеток. Выдвигается также предположение о том, что дофамин играет энергетическую роль в неостриатуме: через цАМФ он обеспечивает распад гликогена.
Помимо теоретического интереса в изучении медиаторной и метаболической функции дофамина, особое значение приобретает участие дофамина в патологии. Было установлено, что у больных с двигательными расстройствами резко падает концентрация дофамина в обоих ядрах неостриатума – хвостатом и скорлупе.
Стриаталамические связи. Неостриатум не имеет отчетливо выраженных моносинаптических связей с корой больших полушарий и с таламусом. Неостриатум осуществляет физиологическую связь с корой большого мозга и таламусом опосредованно, через бледный шар, который выступает в этом случае как неспецифическое ядро, как посредник в эфферентной импульсации хвостатого ядра и скорлупы. Постулируется замкнутый круг импульсации: неостриатум – паллидум – таламус – лобные доли – неостриатум. Этот круг носит название «каудатная петля». Ему придают большое значение в интеграции нервных процессов на высших уровнях мозга, в генезе синхронной активности коры, в регуляции сна и бодрствования.
Кортикостриарные связи. Сейчас доказано, что почти от всех полей коры к хвостатому ядру и скорлупе конвергируют прямые волокна в составе внутренней капсулы и подмозолистого пучка. Наибольшее количество волокон идет к скорлупе и хвостатому ядру от передних отделов коры. Кортикостриарные волокна отличаются пространственной организацией. Топографически это проявляется в том, что передние области коры больших полушарий представлены в головке хвостатого ядра, а задние – в каудальном отделе хвостатого ядра (рис. 2.8).
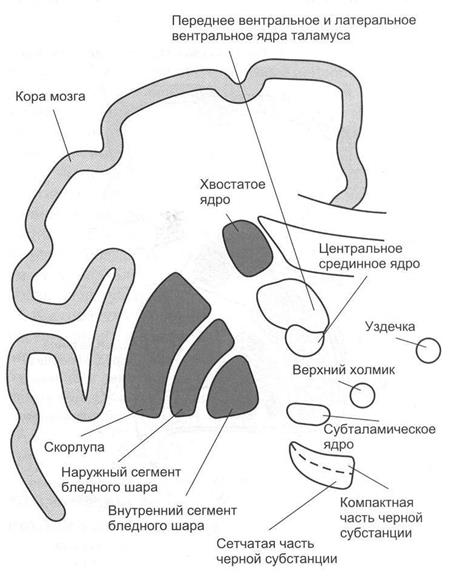
Рис. 2.8. Базальные ганглии и структуры, связанные с ними
Функции базальных ганглиев. Этот комплекс ядер довольно широко включается в интегративную деятельность центральной нервной системы. Они играют определенную роль в ориентации животных в пространстве, запуске двигательного обеспечения пищевой мотивации, регуляции цикла бодрствование – сон. Неостриатум, паллидум, клауструм входят в программу осуществления условного рефлекса. Базальные ганглии и мозжечок являются равнозначными центрами, участвующими в программировании движений. Базальные ганглии могут иметь особое значение для осуществления стереотипных «червеобразных движений». Кроме того, каждая из структур обладает своими функциональными особенностями при вкладе в организацию движения. Неостриатум принимает участие в регуляции медленных движений, в которых преобладает тонический компонент. Паллидум дифференцирует характер движений: так, активность его нейронов у обезьян изменялась под влиянием толкательных движений, но эти же нейроны не реагировали на пронационные движения. Активность клауструма (у кошек) резко учащалась при болевых раздражениях. Отмечено также, что функциональные проявления базальных ганглиев определяются не столько связями отдельных ядер между собой, сколько связями каждого из них с другими структурами центральной нервной системы. Из этих структур наибольшее значение имеют неокортекс, неспецифические ядра таламуса, субталамическое ядро, черная субстанция, гипоталамус. На этом основании в настоящее время выделяют ряд функциональных петель базальных ганглиев.
Скелетомоторная петля имеет входы от премоторной, моторной и соматосенсорной областей коры мозга. Основной поток информации идет через скорлупу, внутреннюю часть бледного шара или каудолатеральную область ретикулярной формации черной субстанции, затем через двигательные ядра таламуса и назад к шестому слою коры больших полушарий.
При регистрации активности индивидуальных клеток скорлупы и бледного шара у обезьян, которые были обучены стандартным движениям, обнаружены четкие корреляции между этими движениями и активностью определенных нейронов. Наблюдается четкая топографическая организация: активность нейронов строго определенной области базальных ганглиев всегда соответствует специфическим движениям конкретных частей тела. Кроме того, во многих случаях наблюдается корреляция с особыми параметрами движения: силой, амплитудой или направлением движения. Регистрация активности клеток показала, что путь от стриатума через латеральную область ретикулярной формации черной субстанции управляет главным образом движением лица и рта.
Окуломоторная (глазодвигательная) петля специализируется, вероятно, на регуляции движения глаз. Входные сигналы поступают от областей коры, контролирующих направление взгляда: фронтального глазного поля (поле 8) и каудальной части поля 7 теменной коры. Затем путь продолжается через хвостатое тело к дорсомедиальному сектору внутренней части бледного шара или к вентролатеральной области ретикулярной части черной субстанции. Затем идут связи к ядрам таламуса, которые дают проекции к фронтальному глазному полю. Аксоны нейронов сетчатой части черной субстанции раздваиваются, и одна ветвь идет к верхнему двухолмию среднего мозга, которое связано с движением глаз. Наблюдается положительная корреляция между активностью этих нейронов и саккадами (резкий перевод взгляда с одной точки на другую). Частота импульсации резко падает перед саккадой, что обусловлено тормозной стрианигральной связью (связью полосатого тела с черной субстанцией). Такое отключение тормозного выхода черной субстанции ведет к фазической активности таламуса или верхнего двухолмия. О полном пространственном разделении скелетомоторной и окуломоторной петель свидетельствует корреляция нейронной активности ретикулярной части черной субстанции с движениями либо глаз, либо рта, но никогда с теми и другими одновременно.
К настоящему времени накоплены анатомические данные о существовании ряда «сложных петель», которые начинаются и заканчиваются в лобных ассоциативных областях коры (дорсолатеральной, префронтальной, латеральной орбитофронтальной, передней поясной), пройдя через ассоциативные ядра таламуса. В ходе филогенеза значительно возрастают размеры и значение корковых структур, стриатума и таламуса, участвующих в сложных петлях, так что у человека они становятся более обширными, чем двигательные. Однако функции сложных петель экспериментально еще не исследованы.
Медиаторная система базальных ганглиев. Прохождение информации в описанных выше множественных параллельных трансстриальных функциональных петлях может облегчаться или подавляться модулирующими системами. Описано несколько модулирующих систем. Особого внимания среди них заслуживает дофаминергическая система. Дофаминергические нигростриальные пути (черная субстанция – полосатое тело) начинаются в сетчатой части черной субстанции. Содержащие дофамин нейроны обнаружены также поодиночке или группами вне черной субстанции, но поблизости от нее.
Очень тонкие дофаминергические аксоны сильно ветвятся, образуя по всему стриатуму относительно диффузную сеть. Вдоль этих волокон находится множество небольших, заметных в световой микроскоп утолщений, называемых варикозами. На электронных микрофотографиях они идентифицируются как пресинаптические элементы. У нейронов сетчатой части черной субстанции довольно регулярная импульсация с частотой 1 Гц. Таким образом, каждую секунду импульс одной дофаминергической клетки вызывает высвобождение дофамина в многочисленных рассеянных по полосатому телу синапсах.
Из-за своего диффузного строения дофаминергическая система не передает детализированную, топографически организованную информацию. Поэтому ее рассматривают как своего рода «ирригационную систему», модулирующую передачу информации по главному каналу. Так, было показано, что высвобождаемый в полосатом теле дофамин модулирует дофаминергическую кортикостриальную передачу (кора больших полушарий – полосатое тело). Восходящие дофаминергические волокна от среднего мозга направляются не только к стриатуму, но и к лимбическим структурам, к префронтальной коре.
Аналогичное модулирующее влияние на базальные ганглии, возможно, оказывают серотонинергические волокна от ядер шва, норадренергические от голубого пятна,а также волокна с неизвестным медиатором от интраламинарных ядер таламуса и от миндалины; все они идут к полосатому телу. Кроме того, в базальных ганглиях содержится множество местных нейронов (интернейронов), модулирующих поток информации в трансстриатных петлях. К ним относятся холинергические нейроны полосатого тела и различные пептидергические нейроны.
В течение длительного времени полосатое тело рассматривали как крупную однородную массу клеток, и лишь недавно была обнаружена его модульная организация. Окончания двух обширных систем афферентных волокон от коры больших полушарий и от ламинарных ядер таламуса образует здесь небольшие четко ограниченные центры. Анатомические эксперименты с дифференциальным окрашиванием волокон, относящихся к разным системам, показали, что в хвостатом ядре перемешаны скопления нервных окончаний от лобной и височной ассоциативной коры. Гистохимические методы дают аналогичную картину: разные медиаторы (глутамат, ГАМК, ацетилхолин, различные пептиды) обнаруживаются в пределах мелких, четко очерченных участков. Сейчас эти центры считаются независимыми компартментами, или микромодулями. Удалось проследить топографическую организацию в виде продольных колонок, идущих через весь стриатум. Таким же образом организованы проекции лобной и височной ассоциативной коры. С помощью микроэлектродного тестирования выявлены соматотопические продольные колонки, относящиеся к скелетомоторной петле. Например, в колонке верхней конечности, вероятно, собираются сигналы от премоторной, моторной и соматосенсорной областей коры. Нейроны в такой колонке объединены по сходству их соматотопических свойств.
Дата добавления: 2016-07-27; просмотров: 4877;
Поиск по сайту
Узнать еще
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- II. Митохондрии (строение и функции)
- II. Признаки, ресурсы и функции власти.
- III. Функции отдела по делам ГОЧС и ВМР
- III. Функции политологии. Возрастание роли политических знаний в жизни общества.
- Table 1. Функции ГИС для лесного хозяйства и лесоустройства
- А.Н.Леонтьев ПВ – это профессиональное общение педагога, имеющее определенные функции, направленное на оптимизацию учебной деятельности
- Абсолютная и относительная погрешности функции

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
