Основные черты популяции как генетического резерва вида
Как следует из определения, для существления популяции необходимы три условия: панмиксия (т.е. возможность свободного скрещивания особи с любой другой этого же вида); отсутствие изоляционных барьеров, нарушающих панмиксию; длительное существование на одной территории.
Панмиксия ззаключается в случайном скрещивании родительских пар. Степень панмиксии внутри популяции должна быть выше, чем между популяциями. Эта разница в степени влияния фактора изоляции между особями и определяет обособление групп особей, именуемых популяциями. Однако внутри популяции неизбежно выделяются особи, скрещивания между которыми носит избирательный характер. Причины такой избирательности в лесных популяциях могут быть как чисто механические, так и генетические. Например, местоположение дерева может лимитировать в большей или меньшей степени как разлёт, так и попадание на женские цветки пыльцы с тех или иных деревьев. Возможна также конкуренция попавших на рыльце цветка пыльцевых зёрен. Причина такой неравноценной возможности у пыльцевых зёрен носит, несомненно, генетический характер. Наконец, генетически обусловлен признак раннего и позднего вступления в фазу цветения у различных деревьев. И всё-таки в популяции в целом влияние факторов, нарушающих панмиксию, не должно быть значительным, так как главная роль панмиксии – обеспечить возможность особям, входящим в популяцию, свободно обмениваться генами, т.е. создать возможно большее число вариаций генотипов предполагаемого видом «рабочего» материала естественному отбору. В популяции, где возможно максимальное перемешивание (комбинация) аллелей, естественный обор оставляет наиболее приспособленные формы организмов. В связи с этим именно панмиксия определяет в первую очередь целостность популяции, как единицы эволюции.
Изоляция нарушает свободную панмиксию внутри популяции, а также ограничивает возможности скрещивания между особями, принадлежащими к разным популяциям. Различают три типа нарушения панмиксии внутри популяции.
Ассортивное скрещивание – избирательность, при которой особи с одинаковыми генотипами образуют пары чаще, чем при случайном скрещивании. К положительному ассортивному скрещиванию может проводить дизруптивный отбор т.е. отбор генотипов с крайними проявлениями признака. Важнейшим результатом ассортивного скрещивания оказывается изменение частот генотипов.
Инбридинг возможен при повышенной частоте скрещивания между особями, относящимися к одной родственной группе. В этом случае идёт гомоготизация по всем локусам.
Селективное скрещивание – особи определённых генотипов чаще по сравнению с особями других генотипов вступают в любые скрещивания. В лесных популяциях при таком типе скрещивания отдельные деревья цветут регулярнее и обильнее, чем другие окружающие их деревья. Селективное скрещивание ведёт к изменению в популяциях не только частоты генотипов, но и генов.
Различают два типа межпопуляционной изоляции: территориальную и биологическую. Изоляция – явление, обратное миграции. Чтобы популяции генетически дифференцировались, необходима известная степень изоляции. С другой стороны, длительный обмен генами нейтрализует эффект отбора.
При территориальной, или географической, изоляции популяции разделены естественным барьером – огромным пространством или горами. Виды, занимающие аридные горные области, имеют сильно разорванные ареалы. Популяции в отдельных горных районов репродуктивно изолированы и тем самым дифференцируются генетически в результате отбора или случайных процессов. Примером разрыва ареалов могут служить островные сосновые боры на южном пределе ареала, берёзовые колки в степной зоне.
Имеются данные, указывающие на очень большой радиус эффективного разлёта пыльцы в пределах леса. Для дуба, например, этот радиус составляет несколько сотен метров, а для сосны от 12 – 20 до 200 м., кедр – 250, ель – до 300, ильм – 2000 м. Семена сосен, елей, пихт, клёнов, ясеней, лип остаются в пределах нескольких сотен метров от деревьев, с которых они падают. Семена берёзы по насту распространяются на несколько сотен метров. У тополей, ив могут лететь на расстояние 0,5 км и более. Семена и дубов, грецких орехов, кедра очень тяжёлые и рассеиваться только на несколько метров от дерева. Могут разноситься белками, кедровкой на значительное расстояние. И всё же среднее расстояние рассеивания их мало, поскольку большую часть семян белки съедают. Некоторые деревья – можжевельники, вишни, сливы, боярышник, малина поедаются птицами и семена разносятся на значительное расстояние от родительских деревьев.
Биологическая изоляция у деревьев чаще всего выражается в различных сроках цветения, обусловленных климатическими факторами. Мутации признака раннего и позднего цветения могут привести к репродуктивно изолированным популяциям. Но если такие мутации произошли и части популяции начали мигрировать в противоположных направлениях, обор может фиксировать гены раннего цветения в одной части, а гены позднего цветения – в другой. Через некоторое количество поколений дифференциация может стать достаточно большой, чтобы предотвратить скрещивание, если субпопуляции опять сольются. Такой предположительный путь возникновения различий во времени цветения между клёнами серебристым (ранний) и красным (поздний). Если сохранить пыльцу, скрестить эти виды искусственно совсем нетрудно. В природе, занимая один и тот же ареал, не скрещивается.
Хозяйственные виды в большинстве своём имеют непрерывное распространение, по крайней мере на большей части ареалов. Вместе с пыльцой или семенами гены могут мигрировать от одного конца к другому этого непрерывного ареала и таким образом предотвращать генетическую дифференциацию.
Райт (1946) разработал теоретическую основу для расчётов влияния изоляции расстоянием. Он ввёл понятие «соседства» – самой большой субпопуляции, внутри которой происходит случайное опыление. «Соседство» – это полоса длиной 2s (p)1/2 (пыльца и семена распространяются одинаково) или s (p)1/2 (распространяется только пыльца). Он установил, что большинство растущих в умеренной зоне древесных видов, имеют средние величины «соседства» 25 – 200 деревьев, что приводит к небольшой или средней локальной генетической дифференциации в пределах 50 – 100 км.
В1908 г. была предложена схема-формула распределения генотипов в популяции, вошедшая в науку под названием закона Харди — Вайнберга. Закон кажется интуитивным и слишком простым, чтобы нуждаться в объяснении. Теорема, используемая для доказательства этого, интересна тем, что даёт простой пример применения математики и селекции. Этот закон гласит: в популяции, в которой осуществляется панмиксия, отсутствует мутационный процесс, отсутствует давление отбора и которая бесконечно велика, сохраняется неизменным равновесие между частотами генотипов, определяемых диаллельным геном А и а, которое выражается следующим образом: р2 (АА) : 2 pq (Аа) : q2(aа)= (p + q)2 =1, где р, q – частота генотипов в популяции. Эта формула показывает численное соотношение генотипов в популяции для одного моногенного признака. И это соотношение сохраняется в популяции из поколения в поколение. Кроме того, из формулы следует, что гетерозиготные генотипы более выгодны для популяции: ведь даже самый редкий аллель будет распространен в популяции в форме гетерозиготы; для малых значений q (если q частота редкого аллеля) q2< 2pq. Эту особенность популяции отметил еще в 1926 г. С. С. Четвериков [1929]: «Видовое сообщество (т. е. популяция) в условиях свободного скрещивания представляет собой устойчивый агрегат, внутри которого в самих условиях свободного скрещивания заложен аппарат стабилизации численных отношений, составляющих его аллеломорфных пар». Для процессов, создающих устойчивость популяции к воздействиям внешней среды, М. Лернером [Lerner, 91] был предложен термин — генетический гомеостаз. Он определил его как процесс, обеспечивающий способность популяции сохранять свою генетическую структуру в ответ на воздействие факторов внешней среды.
Пример. Родительское поколение образует гаметы Аи а в отношении p : q. Последние вF1 образуют деревья АА, Аа и аа, частота встречаемости которых соответствует биномиальному распределению p2 : 2pq : q2. Деревья F1 образуют гаметы следующим образом: деревья АА даютp2 гамет А; деревья Аа – pq гамет А иpq гамет а; деревья аа – q2 гамет а. Все деревья вместе дают (p2 + pq) гамет А и (pq + q2) гамет а. Таким образом, отношение гамет А к гаметам а в поколении F1 равно (p2 + pq) : (q2 + pq) = p(p+q) : q(p + q) = p : q, как в исходной генерации.
ГЕНЕТИЧЕСКИЕ ПРОЦЕССЫ В ПОПУЛЯЦИЯХ
В своей книге “Происхождение видов” Ч. Дарвин в 1859 году впервые объяснил процесс видообразования, вскрыв факторы эволюции:
изменчивость, наследственность и отбор.
Вид можно рассматривать, как исторически сложившуюся совокупность организмов, занимающих определенный ареал обитания и характеризующихся общностью происхождения, сходной системой адаптации к условиям среды и воспроизведением в поколениях основных черт и признаков вида. Особи, составляющие вид, в отношении наследственных свойств не являются однородными. Каждый организм, обладая общими характерными для вида чертами, имеет и свои индивидуальные генотипические особенности. Всю генетическую информацию вида, то есть полный набор генов, сложившийся в процессе его эволюции, называют генофондом вида.
Особи вида расселяются на занимаемой им территории не равномерно, образуяпопуляции.
По определению Л.Ф. Правдива,популяция представляет собой “... группы свободно скрещивающихся или потенциально способных к скрещиванию организмов особей одного вида. населяющих в течение достаточно длительного времени (большого числа поколений) определенную территорию и обнаруживающих пространственно-временные взаимоотношения”.Популяция - это наименьшая элементарная единица эволюции. Сам вид занимает большой ареал, содержит многообразие форм и является громоздкой системой для анализа генетических процессов, происходящих в нем, а отдельный организм (особь) содержит лишь незначительную часть генетической информации вида, которая утрачивается со смертью этого организма.
Исследования по экспериментальному изучению популяций начались с выхода в 1926 году работы С. С. Четверикова “О некоторых моментах эволюционного процесса с точки зрения современной генетики”, и положили начало развитию в нашей стране популяционной генетики. В нее входят как частные вопросы изучения отдельных видов, так и теоретические разработки, на базе которых можно проводить расчеты в популяциях всех видов животных, растений и человека.
Учение о генетических процессах в популяции особенно важно для лесоводов, так как включает в себя вопросы динамики, численности вида, систематики и таксономии, генетического анализа. Жизнь популяции с генетической точки зрения - это динамика частот генов и генотипов в пространстве и времени. Распределение этих частот, их изменение могут происходить под действием отбора. Чтобы изучить эти закономерности, английский математик Д. Харди и немецкий биолог Г. Вайнберг в 1908 году независимо друг от друга создали модель идеальной популяции, где нет отбора, мутаций, миграций, дрейфа генов, и провели математический анализ распределения генов в популяции. Они пришли к выводу, что соотношение генотипов в популяциях, при соблюдении вышеперечисленных условий, сохраняется из поколения в поколение (закон Харди - Вайнберга).
Рассмотрим популяцию, состоящую из очень большого числа особей, в которой скрещивание происходит случайным образом. Допустим, что частота встречаемости гена А равнар, его рецессивной аллели а равна q, тогда р + q = 1 . Гаметы, несущие эти аллели, соединяются между собой случайно, и зиготы, получающиеся в результате такого слияния гамет, будут иметь генотипы и частоты, представленные в табл.
Частота генотипов в популяции
| ♀ ♂ | А Частота p | а частота q |
| А | АА | Аа |
| Частота p2 | P | Pq |
| A Частота q У | Аa Pq | Аa q2 |
| То есть |
| Генотип АА Аа Аа |
| Частота Р2 2pq q2 |
тогда р2 АА + 2pqAa + q2 aa= 1.
В следующем поколении гомозиготные генотипы дадут гаметы только одного типа (АА ? А; А;аа > а, а), а гетерозиготные -равное число гамет одного и другого типа (Аа> А;а).
В идеальной популяции концентрации гамет с аллелем А и аллелем а, которые дают следующие поколения, будет иметь такой вид:
от родителей с аллелем А
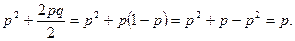
от родителей с аллелем а
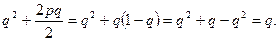
Таким образом, в идеальной популяции концентрации генов не изменяются по поколениям.
Такие идеальные популяции находятся в равновесном состоянии. В природе таких идеальных состояний не существует, так как всегда в популяции происходят миграция, дрейф генов, возникают мутации и происходит отбор. Однако, как заметил С.С. Четвериков еще в 1926 году, "Видовое сообщество (то есть популяции) в условиях свободного скрещивания представляет собой устойчивый агрегат, внутри которого, в самих условиях свободного скрещивания, заложен аппарат стабилизации численных отношений составляющих его аллеломорфных пар". В 1929 году С. Дайвер, изучая полиморфизм раковины у Ceparea nemoralis, показал, что соотношение полосатых и бесполосых раковин в популяциях сохранилось неизменным с плейстоцена до наших дней.
Ю.П. Алтухов и Б.А. Калабушкин изучили соотношение бесполосых и полосатых раковин у брюхоногого моллюска Littorina squalida в современном и ископаемом материале в лагуне Буссе на Южном Сахалине. За 5-6 тысяч лег сменилось 2 000 поколений моллюсков, но сдвигов концентрации аллелей окраски раковин не произошло.
Ю.Г. Рычков показал, что за время от неолита до наших дней, то есть за сотни поколений, при суммарном анализе генных частот в популяциях североазиатской расы монголоидов не наблюдается сколько-нибудь заметных сдвигов этой популяции в целом.
А.В. Чудный на сосне обыкновенной показал, что в сеянцах уровень гетерозиготности по ряда аллелей значительно ниже, чем у деревьев более старшего возраста. С увеличением возраста подроста и до выхода его в первый ярус деревьев, уровень гомозиготности снижается, а гетерозиготности возрастает, достигая постепенно уровня предыдущего поколения. Следовательно, естественный отбор сохраняет в определенном состоянии популяцию в целом, не допуская возможности оставить потомство особям, которые могут нарушить оптимальное соотношение аллелей для данной популяции. Это явление было названогенетическим гомеостазом, то естьравновесием, способностью сохранять популяцией своей генетической структуры в ответ на воздействие факторов внешней среды.
Закон Харди-Вайнберга позволяет рассчитать соотношение генотипов при разных концентрациях аллелей. Анализ распределения фенотипов позволяет в любой реальной системе определить концентрацию генов.
Например, у людей частота появления альбиносов в среднем составляет около 1 на 20 000 (0,00005). Люди-альбиносы являются гомозиготными по рецессивному аллелю а. Среди “нормальных” людей имеются особи, гомозиготные по нормальному аллелю, то есть с генотипом АА, и особи, гетерозиготные - с генотипом Аа. Можно рассчитать концентрацию генов А и а и узнать количество гетерозигот Аа, часть из которых, вступая в брак между собой, рождает детей альбиносов аа. Из указанного выше числа альбиносов в популяциях следует, что концентрация аллеля а равна
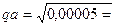 0,007,
0,007,
концентрация аллеля А
рА= 1-0,007 =0,993.
Частота доминантных гомозигот АА составляет
Р2 АА = 0,9932 = 0,98605, то есть 98,6 %.
Частота гетерозигот Аа равна
2pqAa = 0,01390,то есть 1,4%.
Частота гомозигот аа равна
qгaa = 0,0072 = 0,00005, то есть 0,0056 %.
Проведенные расчеты показали, что хотя альбиносыаа встречаются редко - 1 : 20 000 особей, вместе с тем, частота гетерозиготАа равна 1 на 77. То есть, даже при малом числе гомозигот, число гетерозигот в популяциях относительно велико. При родственных браках в семьях, где есть рецессивный аллель, вероятность встреч гетерозигот в качестве отца и матери возрастает, что ведет к выщеплению разнообразных рецессивных генов. Это является причиной более частого рождения наследственно отягощенных детей в браках, где отец и мать являются близкими родственниками.
Популяционные исследования древесных пород были начаты в конце 50-х годов 20-го столетия. Генетический анализ лесных древесных пород по потомству затруднен, так как требует значительного времени для смены поколений. Кроме того, большинство методов генетического анализа разработано на моногенных признаках, которых у древесных видов пока выявлено не так уж много: монотерпены у сосен, пираксидазы у ели. Структурный ген имеет определенную нуклеотидную последовательность, которая отражается в аминокислотном составе детерминирующего им белка (фермента). Ферменты занимают основное положение в метаболизме веществ в клетке. У ели таким ферментом являются изоферменты эстераз (изоэнзимы). При помощи электрофореза получают спектр изоферментов, которые имеют различную величину полос и по их сходству и различию изучают генотипический состав популяции. Семена хвойных древесных пород - удобные объекты для генетических исследований изоферментов. Оболочка семени формируется из диплоидной ткани материнского растения; зародыш - из диплоидной ткани, включающей гомологичные хромосомы отца и матери; эндосперм -из гаплидной ткани материнского организма. Таким образом, анализируя одновременно хвою дерева и эндосперм его семян, можно обнаружить спектр ферментов не только диплоидной ткани, но и каждого из двух составляющих его гаплоидных наборов в отдельности. Наличие двух типов спектров у гаметофитов одного и того же дерева позволяет сделать заключение о гетерозиготности дерева по этому признаку, а обнаружение только одного типа - о его гомозиготности. Впервые Бертельс при скрещивании ели европейской исследовал гибрид и родителей по эстеразам. Он выяснил, что родители отличаются по характеру эстераз, а потомство имеет их гибридную комбинацию. П. Ферет установил корреляцию изоэнзимов сеянцев с изоэнзимами родителей при анализе потомства от самоопыления и контролируемого опыления.
Показатель частоты локусов изоэнзимов пероксидозы былиспользован В.М. Роне в качестве критерия генетического родства в пределах популяции ели обыкновенной.
Л.Я. Полозова и Г.А. Абатурова провели сравнение двух популяций сосны обыкновенной, произрастающих на западе и севере ареала европейского подвида сосны обыкновенной. Сравнение проводили по морфологическим признакам и изоферментам эстераз в эндосперме семян. В итоге было показано, что литовская популяция сосны обыкновенной отличалась от ленинградской не только по морфологическим признакам -окраске шишек и форме апофиза, но и по изоэнзимам.
Наиболее глубокие изучения монотерпенов сосны обыкновенной были проведены А.В. Чудным. Им было установлено, что в терпенных маслах имеются а-пинен, ?-пинен и ?3-карен, синтез которых контролируется единичными генами. При этом по соотношению а-пиена, Я-пиена и ?3-карена было выделено 4 типа биосинтеза монотерпенов, каждому из которых соответствуют определенные генотипы. Наличие большого количества ?3-карена и ?-пинена или их малое содержание обусловлено у каждой особи одной парой аллелей. Обозначим доминантную аллель но ?3-карену знаком С, рецессивную - с, по -пинену, соответственно В и b. Тогда особи, получившие от отца и матери доминантные гаметыСВ, будут синтезировать большее количество ?3-карена и Я-пинена, особи с рецессивными аллелями будут продуцировать терпентинное масло с низким содержанием ?3-карена и Я-пинена. Генотипы ССВВ и СсВbдают фенотипы с высоким содержанием ?3-карена и Я-пинена, ccbb - с низким содержанием этих компонентов.
Эмпирическим путем были установлены границы между доминантными томозиготамиСС и ВВ и гетерозиготамиСс и Вb, что было подтверждено и анализом потомства при скрещивании.
По анализу терпенных масел можно определить генетический состав популяции сосны и, если она отвечает определенным требованиям, создать искусственное насаждение, которое будет таким же, как модельное. Например, А.В. Чудный рассчитал оптимальный состав генотипов сосны обыкновенной для ЛСП. Им были определены генотипы сосны в местном насаждении в спелом насаждении и молодняках (табл.).
Таблица . Соотношение генотипов сосны
| Генотипы | Частота | Относительная выживаемость | Степень адаптации | Доля участия генотипа на плантации, % | |
| Молодняк, % | Спелое насаждение,% | ||||
| ЬЬсс | 33,2 | 40,0 | 1,2 | 0,81 | 22.5 |
| ЬЬСс | 17,6 | 26,0 | 1,48 | 1,0 | 27,6 |
| ЬЬСС | 43,2 | 28,0 | 0,67 | 0,45 | 12,5 |
| ВВсс | 4,0 | 4,0 | 1,0 | 0,67 | 18,7 |
| ВЬСс | 2,0 | 2.0 | 1,0 | 18,7 |
х=3,6 100%
Относительная выживаемость определяется отношением частот генотипов в спелых насаждениях к аналогичным генотипам в молодняках. Например, относительная выживаемость генотипа bbcc равна 1,2 (40,0:33,2).
По относительной выживаемости определяется степень адаптации генотипов. За единицу принимается наибольшее значение генотипа по относительной выживаемости. Для приведенного примера за единицу принимается относительная выживаемость генотипаbbСс, а степень адаптации остальных генотипов рассчитывается в долях от единицы. Например, степень адаптации генотипаbbcс рассчитывается следующим
образом: 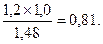
Общий адаптационный коэффициент (3,6) - это сумма степени адаптации всех генотипов, которая принимается за 100 %. От него рассчитывается доля участия каждого генотипа в насаждении. Например,
доля участия в насаждении генотипа bbcc 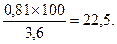
Таким образом, чтобы воссоздать исходную популяцию сосны обыкновенной, нужно создать лесосеменную плантацию, на которой доля участия генотипов должна составлять соответствующий процент.
Вопросы для самопроверки
1. Дайте определение вида и популяции.
2. Почему популяцию считают элементарной единицей эволюции?
3. Что такое идеальная популяция?
4. Дайте формулировку закона Харди-Вайнберга. Приведите общую формулу этого закона.
5. Докажите, что частота генотипов в популяции не изменяетсясовременем.
6. Объясните понятие гомеостаза.
7. Как закон Харди-Вйнберга объясняет увеличение гомозигот при близкородственных браках?
8. Значение популяционных исследований для древесных пород. Приведите пример.
9. При помощи какого анализа можно определить состав изоферментов древесных растений? Приведите примеры.
Дата добавления: 2017-09-01; просмотров: 3148;
Поиск по сайту
Узнать еще
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- Cущность организации и ее основные признаки
- I. Демократия как тип политического режима.
- I. Назначение унифицированных газозарядных станций и основные тактико-технические требования, предъявляемые к ним.
- I. ОСНОВНЫЕ ПОЛОЖЕНИЯ
- I. Политический режим: понятие, сущность и основные типы.
- I.2. Основные категории водопотребления промышленных предприятий и их особенности
- II. Агрессия как проявление побуждения

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
