Изменчивость, обусловленная взаимодействием между генотипом и средой
Ключевое значение при исследовании изменчивости имеет взаимодеиствие “генотип — среда”. Этот термин используется, чтобы представить изменчивость тех или иных генотипов, растущих в различных условиях среды. Такое взаимодействие должно быть изучено, если желают получить максимум прогресса от селекции. Слишком часто при селекции растений группы семей испытывают в одном единственном виде условий местопроизрастания. Полученные при этом результаты затем экстраполируют на другие условия. Фактически эти же генотипы при выращивании в других условиях могут дать совершенно иные результаты.
Ввиду этого, коммерческие сорта лесных древесных пород, как и сельскохозяйственные растения, должны изучаться в нескольких существенно различающихся условиях местопроизрастания.
Важно не путать настоящее взаимодействие “генотип — среда” с npостым ответом на различия условий среды, где относительная разница между испытуемыми семьями остается по существу той же самой, если для средние показатели семей в различных условиях изменяются значительно. Например, средняя высота семьи А составляет 20,0 м в одних условия и только 17,0 м в других, в то время как семья Б имеет 21,5 м в первых условиях и 18,0 м в других. Различие в высотах между одними и теми же cемьями в двух различных условиях есть только ответ на различия в условиях выращивания, но здесь нет никакого взаимодействия “генотип — среда”, поскольку ранги семей А и Б не изменились существенно в этих двух разных условиях. Общераспространенной ошибкой является представление различий от одного местоположения к другому как взаимодействие “генотип — среда”.
Настоящее взаимодействие “генотип — среда” встречается при широком варьировании условий местопроизрастания. Если при этом происходит смена рангов роста отдельных генотипов, то можно говорить о наличии взаимодействия “генотип — среда”. Например, семья А в условиях имеет среднюю высоту 19,0 м, а в условиях У— 21,5 м, в то же время семья Б в условиях Х имеет высоту 22,0 м, а в условиях У— только 20,0 м. Налицо смена рангов высот семей А и Б в разных условиях местопроизрастания. Изменчивость, обусловленная наличием взаимодействия “генотип — среда”, может существенно изменить наши представления о ценности тех или иных генотипов и возможности их хозяйственного использования.
В зависимости от поведения тех или иных генотипов в разных условиях местопроизрастания можно оценить их генетическую стабильность и принимать соответствующие решения по их выращиванию. Более подробно о взаимодействии “генотип — среда” см. также в главе .
Вопросы для самопроверки
/. Что такое биоразнообразие, и какие его уровни Вы можете назвать?
2. Какой уровень биоразнообразия наиболее важен для лесов умеренной зоны и почему?
3. Какими тремя главными причинами обусловлена внутривидовая изменчивость?
4. Какие типы и формы изменчивости древесных растений выделяет С. А. Мамаев?
5. У каких наших лесных древесных пород наиболее изучена внутривидовая изменчивость и кем?
6. Какие термины используют при выделении внутривидовых географических категорий лесных древесных пород? Приведите их определения.
7. Что такое происхождение, географический источник и географическая раса?
8. Чем отличается термин “географический источник” от термина “источниксемян”?
9. Что такое раса и экотип? В чем сходство и различие этих понятий?
10. Что подразумевается под термином “клин”? В чем отличие клина от экотипа?
11. Что такое ландраса? Какие условия отбора необходимо соблюдать для успешного использования ландрас?
12. Какой метод может быть основным у лесоводов для преодоления неблагоприятных экологических воздействий на лесные древесные породы?
13. Назовите основные составляющие генетической изменчивостилесных древесных пород.
14. Какими генетическими составляющими изменчивости обусловлены в большей степени такие характеристики лесных древесных пород, как рост, устойчивость и качество древесины?
15. Каковы последствия самоопыления? Какие варианты этих последствий Вы можете назвать?
16. Что такое взаимодействие “генотип — среда”? В чем различие между простым ответом организма на воздействие окружающей среды и ответом, связанным со взаимодействием “генотип — среда”? Приведите пример.
3.5. ЗАКОН ГОМОЛОГИЧЕСКИХ РЯДОВ В НАСЛЕДСТВЕННОЙ ИЗМЕНЧИВОСТИ Н.И. ВАВИЛОВА
Еще Ч. Дарвин в 1859 году отмечал проявления одинаковых признаков у различных видов растений и животных. Это явление им было названо параллельной изменчивостью. Дарвин пишет: “Под этим термином я разумею, что одинаковые признаки, время от времени, проявляются у некоторых разновидностей или рас, ведущих начало от одного и того же вида и, реже, в потомстве отдаленных видов... Случаи аналогичной изменчивости в отношении их происхождения могут быть подразделены на две категории: во-первых, на случаи, зависящие от неизвестных причин, действующих на одинаковые конституции, а отсюда и варьирующих одинаково, и, во-вторых, на такие случаи, которые обязаны выявлению признаков более или менее отдаленных предков”. До Ч. Дарвина об этом в своих работах упоминают ботаник Ш. Нодэн и энтомолог Уолш. Позднее, Г. де-Фриз в “Мутационной теории” и “Пангенезе” отметил ряды параллельной изменчивости. Г. Осборн, Э. Коп отметили проявление параллельной изменчивости у ископаемых животных. В 1907 году П. Саккардо и Э. Цедербауэр приводят данные по параллелизму у грибов и хвойных.
Частные указания систематиков на закономерности параллелизма в изменчивости в отдельных группах растений и животных, и особенно насекомых, показывают широкую распространенность этого явления.
Детальные исследования изменчивости многих видов, огромное количество новых фактов, собранных на культурных растениях и близких к ним диких родичах, позволили Н.И. Вавилову заново подойти к этой проблеме и свести все известные факты в форму общего закона, которому подчинены все организмы и который должен быть положен в основу систематизации знаний о наследственной изменчивости видов.
В 1920 году Н.И. Вавилов сформулировал закон гомологических рядов в наследственной изменчивости, согласно которому родственные по происхождению виды растений и животныхимеют сходные ряды наследственной изменчивости.
1. Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и виды, тем полнее сходство в рядах их изменчивости.
2. Целые семейства растений характеризуются определенным циклом изменчивости, проходящем через все роды и виды, составляющие семейство.
Вышеуказанные закономерности Н.И. Вавилов выразил формулой закона гомологических радов для видов и родов. Для видов (линеонов) формула имеет следующий вид:
L1 (a+b+c+d+e+f+g+h+i+k+...),
L2(a+b+c+d+e+f+g+h+i+k+...),
L3(a+b+c+d+e+f+g+h+i+k+...),
где L1, L2, L3 - радикал, отличающий виды (линеоны) один от другого;
а, Ь, с ... – различные варьирующие признаки - форма чешуи, листьев, стеблей, окраска листьев, хвои, форма кроны и так далее. Каждый из этих признаков сам по себе сложен и может быть соответственным образом разделен на большее или меньшее число морфологических и физиологических единиц: a1, а2, а3 и так далее.
Для родов формула имеет следующий вид:
g1 (a+b+c+d+e+f+g+...),
G2 (a+b+c+d+e+f+g+...),
Сз (a+b+c+d+e+f+g+...),
где G1, G2, С3 - радикал отличающий роды один от другого;
а, Ь, с,... - варьирующие признаки.
Так как многие роды включают значительное число видов, то более правильное представление о роде дает следующая формула;
G1 (a+b+c+d+e+f+g+h+...)(L1+L2+L3+L4+L5).
Например, обозначим радикал рода пихта (Abies Mill.) через G1, радикал вида пихты белой (Abies alba Mill) – L1; радикал вида пихты кавказской (Abies Nordmanniana (Steven) Spach.) – L2; радикал вида пихты сибирской (Abies sibirica Ldb.) — L3; радикал вида пихты бальзамической (Abies balsamea (L.) Mill.) – L4; радикал вида пихты одноцветной (Abies concolor (Gord.) Engelm.) - Ls.
Обозначим радикалы варьирующих признаков различных видов по габитусу (характеру роста, ветвлению и формы кроны) - а, по окраске хвои — b. Различные выражения этих признаков обозначим через a1, а2, а3...; b1, b2, b3 и так далее.
Так, по габитусу у различных видов пихт выделены следующие формы: пирамидальная (f. pyramidalis) — a1; колонновидная (f. columnaris) — a2; плакучая (f. pendula) – а3; прямостоящая (f. erecta) – а4; распростертая (f. prostrata) – а5, шаровидная (f. globosa) –a6; канделябровая (candelabrum) – a7; низкая (f. pumila unuf. папа) – a8.
По окраске хвои: пестрая (f. variegatd) – b1; золотистая (f. aurea) — b2; золотистокончиковая (f. aureo-spicata) - bз; сизая (f. glauca) – b4; белая (f. alba) –b5; изящная (f. elegans) – b6; серебристая (f. argentea) – b7, голубая (f. violacea) – b8.
Тогдаg1 (a1 + a2 + а3 + а4 + а5 + a6+ а7 + a8 + b1 + b2 + b3 + b4 + b5 + b6 + +b7+b8)(L1+L2+L3+L4+L5).
Перемножив между собой содержимое скобок, получаем гомологические ряды наследственной изменчивости перечисленных видов пихт:
L1 a1 +L1a2+L1a3 + L1a4 + L1a5 + L1a6+ L1a7 + L1a8+L1b1 +L1 b2 +L1b3 +L1b4+L1b5+L1b6+ L1b7+L1b8;
L2а1 + L2а2 + L2а3 + ….+L2 b2 +L2 b3 +L2 b4 …
G1 L3а1+L2а2…+L3 а3 …+L3 а7+L3 а8 +L3 b1…+L3 b4+L3 b5+L3 b6…
L4а1+ L4а2…+L4 а5 …+L4 а8+L4 b1 …+L4 b4 …+L4 b7…
L5а1+ L5а2+ L5а3 …+L5 а6 …+L5 b2 …+L5 b7+L5 b8
Жирным курсивом выделены формы, имеющиеся у данного вида. Так, форма пирамидальная обнаружена у двух видов пихт из пяти, колонновидная - у трех; повислая - у четырех; прямостоящая - у одного; распростертая - у одного; шаровидная - у одного; кандедябровая - у одного; низкая - у двух; форма пестрая - у трех; золотистая - у трех; золотистокончиковая - у одного; сизая - у двух; белая - у одного; изящная - у одного; серебристая - у двух и голубая - у одного. Не выделенные формы еще не обнаружены в природе или не выведены искусственным путем.
Сам Н.И. Вавилов закономерности в полиморфизме у растений, образующих гомологические ряды в наследственной изменчивости, сравнивал “... с гомологическими рядами органической химии, например, углеводородами (CH4,, C2H4…). Ряды этих соединений, отличаясь друг от друга, все же характеризуются многими общими свойствами в смысле образования определенных циклов соединений, определенных реакций обмена и соединения”. Он проанализировал параллельную изменчивость не только в пределах отдельных родственных групп, но и в разных семействах, генетически не связанных, даже в разных классах. Так, альбинизм, то есть появление бесхлорофильных растений и полуальбиносов, наблюдаются в самых разнообразных семействах:
сосновые, кипарисовые, барбарисовые, розанные, камнеломковые, самшитовые, кленовые, бересклетовые и другие. Различные виды этих семейств имеют одинаковые формы: пестролистные (f. variegata), серебристая (f. argentea), золотистая (f. ангеа), золотисто-пестрая (f. aигео-variegata), серебристо-пестрая {f. cdba-variegata), золотистоокаймленная (f. aureo-marginatd) и другие.
Существует другое проявление параллельной изменчивости -образование карликовых и гигантских форм -нанизм и гигантизм. Например, у семейств кипарисовые, тисовые, сосновые, буковые, маслинные имеются формы карликовая (f. pygmaea), низкая (f. папа), распростертая (f. prostrates) шаровидная (f. globosa), малорослая {f. pumila), наибольшая (f. maxima), монументальная (f. monumentalis) и другие.
Во многих семействах проявляется тенденция к образованию фасциации - расширение различных органов. Н.И, Вавилов наблюдал это явление у гороха, сои, льна, свеклы, подсолнечника, ячменя, кукурузы, пшеницы, гречихи, тыквы, арбуза и других. В дендрологии известны виды, имеющие фасциации. Например, у видовой нормы ясеня обыкновенного (Fraxinuc excelsior L.) листья непарноперистые, состоят из 7...9 (5...15) листочков, а у однолистной формы (f. diversifolia) листья простые одиночные или отчасти тройчатые.
Н. И. Вавилов отмечает, что карликовость, гигантизм, альбинизм и фасциация проходят по всему растительному миру и проявляются даже в животном мире.
Примеры параллельной изменчивости можно найти в различных семействах и по другим признакам, таким, как плотность соцветия, форма плодов, окраска цветков, плодов и семян, форма ствола и его окраски и многое другое.
Наличие параллелизма в полиморфизме, существование правильностей в дифференциации видов, родов и семейств облегчают изучение разнообразия растений и животных.
Закон гомологических рядов наследственной изменчивости можно сравнить по значимости с периодической системой элементов Д.И. Менделеева. Как отмечал Н.И. Вавилов, закон гомологических рядов позволяет предугадать открытие форм, еще не найденных у данного вида, но уже обнаруженных у другого близкого вида, а также получение искусственно, путем мутаций, инцухта или гибридизации соответствующих форм. Так, на основании этого закона были найдены яровые формы дикого ячменя, сегментированные формы арбуза, формы чечевицы с зелеными семядолями, формы люпина, не содержащие алкалоидов, из которых впоследствии было выведено множество сортов кормового люпина (в России 16 сортов).
В настоящее время с полным основанием можно сказать,что у родственных видов, имеющих общее происхождение, возникают и сходные мутации. Более того, даже у представителей разных классов растений и животных мы встречаем параллелизм - гомологические ряды мутаций по морфологическим, физиологическим и другим признакам и свойствам.
Исходя из закона гомологических рядов, следует принять, что если обнаруживаются спонтанные или индуцированные мутации у одного вида растений или животных, то можно ожидать сходный ряд мутаций и других видов этого рода. То же относится и к более высоким систематическим категориям.
Наиболее вероятное объяснение гомологических рядов наследственной изменчивости сводится к следующему:
1) родственные виды внутри одного рода, роды внутри одного отряда или семейства могли возникнуть посредством отбора форм с различными полезными хромосомными перестройками. В этом случае родственные виды, разошедшиеся в эволюции за счет отбора разных хромосомных перестроек, могли нести гомологичные гены как исходные, так и мутантные;
2) виды могли возникнуть также путем отбора спонтанных полиплоидов, содержащих однородный набор хромосом;
3) закрепление мутаций вне зависимости от родства. Дивергенция видов, идущая на основе этих трех типов следственной изменчивости, обеспечивает общность генетического материала у родственных систематических групп. Если нуклеиновые кислоты (ДНК, РНК) в комплексе с белком являются тем первичным субстратом, который обеспечил с самых ранних этапов эволюции программирование живых систем, то закон гомологических рядов приобретает всеобщее значение, как закон возникновения аналогичных рядов биологических механизмов и процессов, совершающихся в органической природе. Это относится и генетическим механизмам всех живых организмов - деление клеток, механизмы митоза, мейоза, мутирования и т. д. То есть живая природа в процессе эволюции как бы программировалась по одной формуле, независимоот времени происхождения того или иного типа организмов.
Подобное гипотетическое соображение требует подтверждение на основе синтеза многих знаний. Исследователи должны искать не столько частные различия, характеризующие дивергенцию видов, сколько их общие черты, в основе которых лежат аналогичные генетические механизмы.
Вопросы для самопроверки
1. Назовите одинаковое проявление признака “окраска хвои” уразличных видов рода пихта.
2. Назовите одинаковое проявление признака “габитус растения” у различных видов рода пихта.
3. Каково значение закона гомологических рядов наследственной изменчивости сформулированного Н.И. Вавиловым?
4. Как можно объяснить одинаковые мутации в различных семействах?
5. Приведите примеры проявления параллельной изменчивости альбинизма, нанизма, гигантизма, фасциации у различных видов древесных растений.
6- Составьте гомологические ряды для видов рода туя, можжевельник, береза, ель, сосна, липа по признакам: окраска хвои или листьев, габитус растения, форма листовой пластинки.
7. При помощи формулы закона гомологических рядов в наследственной изменчивости установите формы, не найденные в природе у видов перечисленных родов.
ФЕНЕТИКА ПОПУЛЯЦИЙ
Большинство признаков, с которыми работают лесные селекционеры, относятся к количественным - высоты, диаметры, приросты и другие. Они чаше полигенны, и выход на ген-признак затруднен. Н.В. Тимофеев-Ресовский предложил использовать в таком случае термин “фен”. В 1976 году решением Первого всесоюзного совещания по фенетике популяций термин“фен” и “фенетика” были закреплены официально.
Фенетика - это распространение генетических подходов и принципов на формы, генетическое изучение которых затруднено или невозможно. Предметом фенетики является внутривидовая изменчивость, доводимая в конечном итоге до рассмотрения дискретных альтернативных признаков -фенов.
Фен представляет собой дискретный, альтернативный простой признак, отражающий особенности данного генотипа. Элементарность признака не обязательно сопровождается моногенностью и может быть результатом взаимодействия нескольких генов. Кроме этого, неделимый фен может представлять собой коррелятивно сцепленную группу признаков. Большинство работ по лесной селекции проведено на уровне фенов, то есть изучено формовое разнообразие вида по количественным и качественным признакам. Однако чаще эти работы носили описательный характер (цвет коры, стробил, листьев, характер ветвления и пр.), так как выделение фена и подбор генетической модели довольно сложны для лесных популяций. Все популяционные модели базируются на законе Харди-Вайнберга, постулирующем равновесие генных концентраций в панмиктической популяции.
Однако в популяциях могут происходить мутации, миграции, случайный дрейф генов, изоляция, нарушающие генетическое равновесие. Сложность генетических и экологических явлений не дает возможности провести анализ всей системы факторов и ведет к упрощению популяционных моделей. Генетики создали многочисленные способы расчетов и модели, учитывающие эти явления в популяциях.
Интересную модель расчета морф (фенов) предложил Л.А. Животовский (1982). При этом для анализа популяционной изменчивости по полиморфным признакам постулируются следующие требования:
1) небольшие изменения частот распространенных морф, а также появление или исчезновение редких морф мало изменяет значение показателя,
2) показатель не зависит от параметров, дифференциально характеризующих разные морфы (от приспособляемости морф,иххозяйственной ценности и пр.);
3) вклад морф в величину показателя должен быть пропорционален их частотам в популяциях;
4) показатель должен иметь “простую” популяционную интерпретацию и обладать “хорошими” статистическими свойствами.
Внутрипопуляционное разнообразие оценивается показателем 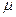 — средним числом морф в популяции, равным
— средним числом морф в популяции, равным
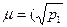 +
+ 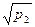 +
+ 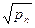 )2
)2
где p1, р2...рn - частоты встречаемости фенов (в сумме равны единице); 
т - наибольшее число фенов в популяции.
При неравномерном распределении числа морф < т. Примономерном распределении 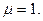
Ошибка S 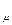 вычисляется по формуле
вычисляется по формуле 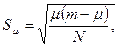
где N- объем выборки.
Доля редких фенов h оценивает структуру разнообразия популяции и определяется по формуле 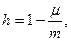
Ошибка Sh вычисляется по формуле 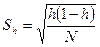
Показатель сходствапопуляций г вычисляется по формуле:
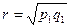 +
+ 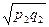 +…+
+…+ 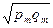 ,
,
где p1... рт - частоты различных морф в первой популяции;
q1... qm - частоты этих морф во второй популяции.
Значение показателя сходства не превышает 1. Если сравниваемые показатели идентичны по числу морф, то r =1. Если нет ни одной общей
морфы, r = 0.
Оценка значимости показателя сходства популяций проводится ври помощи критерия идентичности I по формуле
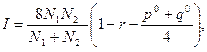
где N1 и N2 - объем выборок популяций,
р° - сумма частот фенов первой выборки, не представленной во второй выборке,
q° - сумма частот фенов второй выборки, не представленной в первой.
Если I превышает табличное значение 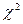 с заданным уровнем значимости и степенями свободы (т - 1), то между выборками есть различие. Если r значительно отличается от 1 (по критерию I), то его выборочная ошибка рассчитывается по формуле
с заданным уровнем значимости и степенями свободы (т - 1), то между выборками есть различие. Если r значительно отличается от 1 (по критерию I), то его выборочная ошибка рассчитывается по формуле
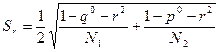
Если же каждая морфа представлена в обеих популяциях, то есть р° = 0; q° = 0, то формула примет вид
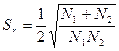 (1-r2)
(1-r2)
Идеальной моделью для фенетического анализа среди древесных пород являются узорчатые формы карельской березы, которая представляет собой, по мнению А.Я. Любавской, самостоятельный вид, имеющий разорванный ареал. Часть этого ареала можно считать группами родственных популяций, состоящих из безузорчатых и узорчатых форм. В основе выделения форм лежит классификация А.Я. Любавской по габитусу и форме ствола, являющимися четкими маркёрами для выделения фенов в популяциях.
Карельская береза (Betula pendula Roth. var. carelica (Mercl.) Hamet-Ahti) является разновидностью березы повислой и отличается от последней не только узорчатой древесиной, но и многими морфологическими признаками. Карельская береза представляет собой кустарник или дерево до 25 м высотой, с утолщениями в виде вздутий, желвачков, хорошо заметных под тонкой берестой молодых деревьев, или крупных почковидных образований и наплывов с перехватами между ними. Крона у древовидных форм карельской березы компактная, вначале яйцевидная, затем округлая, со свисающими концами тонких ветвей. У кустовидных и кустарниковых форм крона раскидистая. Произрастает карельская береза как на бедных каменистых и песчаных почвах, так и на более богатых, без избыточного увлажнения. Кроме этого, она встречается в горных областях, среди скал, каменистых россыпей, в сельгах, урочищах, то есть при резко выраженном рельефе, а также на холмах, склонах оврагов, балках. Растет карельская береза в смешанных насаждениях с участием ольхи серой, березы повислой, осины, сосны обыкновенной и ели европейской, занимая площади от нескольких квадратных метров до 2-3 га.
Являясь светолюбивой породой, карельская береза растет в насаждениях с невысокой полнотой. В смешанных сосново-березовых насаждениях доживает до 65-80-летнего возраста.
Места произрастания карельской березы находятся между 50° и 63° северной широты и 10° и 39° восточной долготы и включают в пределах России Карелию, Костромскую, Владимирскую, Калужскую области. За пределами России она встречается в Белоруссии, странах Балтии, Германии, Словакии, Польше, Норвегии, Швеции, Финляндии.
Красивая текстура древесины, необычное строение ствола привлекало и привлекает к карельской березе многих исследователей. Изучение карельской березы в нашей стране и за границей показало ярко выраженный полиморфизм ее по морфологическим признакам и текстуре древесины. Карельская береза растет деревом второй или третьей велечины, кустарником с белой или темной корой, гладковатой или трещиноватой, с сильно выраженными или незначительными утолщениями, с волнистой, крупно- и мелкоузорчатой древесиной. Исходя из этого, были предложены различные классификации, в основе которых лежало выделение габитуальньк форм. Наиболее удачная классификация А.Я. Любавской, которая выделила 7 основных форм карельской березы - 6 форм с признаками, и одну - без признаков узорчатости. В этой классификации, кроме габитуса, был учтен характер утолщений ствола. Из узорчатых форм четыре относятся к древовидной группе и две - к кустовидной. При этом к формам, близки по габитусу, добавлен индекс “а” и “б” и отмечается характер узорчатости древесины:
1-а - высокоствольная крупноузорчатая форма; 1-6 - высокоствольная шаровидноутолщеная неравномерно-узорчатая форма; 11-а - короткоствольная пятнисто-узорчатая форма; 11-б - короткоствольная лироствольная плотно-узорчатая форма; Ш - кустовидная мелкоузорчатая форма; IV - кустарниковая соединенно-узорчатая форма; V -полнодревесная безузорчатая форма.
В генетический анализ были включены только узорчатые формы, при этом высокоствольные узорчатые формы были объединены в первую форму (1), короткоствольные - во вторую (II). Было обследовано 7 популяций карельской березы: две в Карелии - Заонежский и Спасогубский заказники;
одна в Белоруссии - Телеханский заказник; две в Латвии - Елгавский и Даугавпилсский леспромхозы; одна в Калужской области - Спас-Деменский лесхоз; одна во Владимирской области - Ковровский лесхоз (Шапкин, Погиба, Казанцева, 1996).
Популяционно-генетический анализ проведен для всех выше перечисленных насаждений карельской березы, отличающихся по числу форм и их соотношению (табл.). Популяции карельской березы отличаются друг от друга средним числом фенотипов. Наибольшее формовое разнообразие наблюдается в Телеханском заказнике Белоруссии, среднее число фенотипов 3,66 ± 0,056, представлены все формы. Преобладают особи П формы – p2 = 0,42. Доля редких фенотипов (кустарниковых форм) незначительна - 0,085 ± 0,14.
Наименьшее число фенотипов имеют латвийские популяции:
Елгавская - 1,39 ± 0,017 и Даугавпилсская - 1,98 ± 0,002, но преобладают особи кустовидных форм - рз = 0,96 и 0,60 соответственно. Редким фенотипомв обеих популяциях являются особи высокоствольной формы.
В Заонежском заказнике преобладают особи Ш формы - рз = 0,50, редкий фенотип - IV форма (0,175 ± 0,030); в Спасогубском заказнике преобладают особи I формы –p1 = 0,70; редкий фенотип - П форма (0,16 ± 0,035); в совхозе "Ковровский" преобладают деревья П формы – р2= 0,53; редкий фенотип - Ш форма (0,21 ± 0,027) и в Спас-Деменском лесхозе преобладают особи 1формы –р1 = 6,60; редкий фенотип - 1У формы (0,30 ± 0,026).
Отличие популяций карельской березы по соотношению форм достоверно на 95 и 99 %-ных уровнях значимости.
Популяционно-фенетический анализ имеет значение не только дм теоретических исследований в генетике, но может использоваться и в селекции для решения практических задач.
Таблица. Показатели сходства и критерии идентичности семи популяций
карельской березы
| Сравниваемые выборки | Показатель сходства, r±S, | Критерий идентичности, / |
| |
| 5% | 1% | |||
| 1-2 | 0,98 ± 0,008 | 20,07 | 7,81 | 11,34 |
| 1-3 | 0,91 ±0,013 | 80,65 | 7,81 | 11,34 |
| 1-4 | 0,74 ±0,032 | 203,62 | 7,81 | 11,34 |
| 1-5 | 0,84 ±0,032 | 94,66 | 5,99 | 9,21 |
| 1-6 | 0,27 ±0,071 | 196,32 | 5,99 | 9,21 |
| 1-7 | 0,53 ±0,056 | 49,68 | 5,99 | 9,21 |
| 2-3 | 0,95 ±0,012 | 72,57 | 7.81. | 11,34 |
| 2-4 | 0,77 ± 0,029 | 215,43 | 7,81 | 11,34 |
| 2-5 | 0,96 ±0,015 | 24,96 | 7,81 | 11,34 |
| 2-6 | 0,39 ± 0,056 | 180,79 | 7,81 | 11,34 |
| 2-7 | 0,68 ± 0,053 | 34,99 | 7,81 | 11,34 |
| 3-4 | 0,88 ± 0,015 | 124,29 | 7,81 | 11,34 |
| 3-5 | 0,88 ± 0.023 | 64,90 | 7.81 | 11,34 |
| 3-6 | 0,48 ±0,042 | 140,51 | 7,81 | 11,34 |
| 3-7 | 0,66 ± 0,032 | 32,47 | 7,81 | 11,34 |
| 4-5 | 0,75 ±0,028 | 115,06 | 7,81 | 11,34 |
| 4-6 | 0,75 ± 0,027 | 47,29 | 7,81 | 11,34 |
| 4-7 | 0,74 ± 0,034 | 23,12 | 7,81 | 11,34 |
| 5-6 | 0,56 ± 0,066 | 111,06 | 5,99 | 9,21 |
| 5-7 | 0,84 ±0,051 | 17,58 | 5,99 | 9,21 |
| 6-7 | 0,88 ±0,063 | 13,71 | 3,84 | 6,63 |
Вопросы
1. Что такое фен?
2. Свойства фена.
3. Что такое количественные и качественные признаки?
4. Что такое фенетический анализ популяций?
5. Приведите пример методики расчета фонетического анализа. Какие при этом условия должны быть соблюдены?
4. ВВЕДЕНИЕ В ГЕНЕТИКУ ПОПУЛЯЦИЙ –
Дата добавления: 2017-09-01; просмотров: 2049;
Поиск по сайту
Узнать еще
- II. Степень насыщенности, определяемая природой связи между атомами углерода.
- II. Участие Республики Казахстан в международных договорах
- IV. Классификация обязательств РК по международным договорам
- N в процессе иммунного ответа антигенам 1-го класса принадлежит ведущая роль во взаимодействии между клеткой-эффектором (Т киллеры) и клеткой-«мишенью»
- VI. Примеры мер по реализации международных обязательств
- VII. Пути развития национального экологического законодательства с учётом международных обязательств РК
- А – по капилляру круглого сечения; б – между параллельными пластинками
- А) Два излучателя при разных фазовых соотношениях и расстояниях между ними.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
