Линкёрная ДНК Гистоновый кор
ХИМИЧЕСКАЯ ОРГАНИЗАЦИЯ ХРОМОСОМ.
Химический состав хромосом - ДНК- 40%, гистоновых белков - 40%, негистоновых - 20% немного РНК, липиды, полисахариды, ионы металлов.
Имеется 5 фракций основных гистоновых белков (H1 , H2A , H2B , H3 , H4) и более 100 фракций кислых негистоновых белков,
Функции гистоновых белков: регуляторная (прочно соединяясь с ДНК препятствуют считыванию информации) и структурная (обеспечивают пространственную организацию ДНК в хромосомах, образуя нуклеогистон).
Функции негистоновых белков: среди них ферменты регулирующие процессы:
· синтеза РНК (полимеразы) и процессинга РНК,
· редупликации и репарации ДНК (геликаза, ДНК полимераза, эндонуклеаза, экзонуклеаза, лигаза),
· регуляторная функция, заключающаяся в «запрещении» или «разрешении» считывания информации с молекулы ДНК
ДНК ЭУКАРИОТИЧЕСКИХ КЛЕТОК ПРЕДСТАВЛЕНА СЛЕДУЮЩИМИ ФРАКЦИЯМИ:
а) уникальные последовательности генов 56 % - присутствуют в гаплоидном наборе в единственном числе, образуют основную часть структурных и регуляторных генов,
б) гены со средним числом повторов 8 % - 102 - 104 копий, это структурные гены кодирующие первичную структуру гистонов или нуклеотидов рибосомальных и транспортных РНК,
в) многократно повторяющиеся гены 12 % -106 копий - нетранскрибируемая сателлитная ДНК, играющая роль спейсеров (фрагментов) разделяющих структурные и регуляторные гены ,
г) <блуждающие> структурные гены, положение которых в хромосоме меняется в зависимости от жизненного цикла,
д) молчащие гены они реплицируются, но не транскрибируются, участвуют в обеспечении структурной организации хроматина и в регуляции экспрессии генов,
Хроматин
Хромосомы в интерфазе.В интерфазе нельзя различать отдельные хромосомы. Интерфазные хромосомы слабоспирализованны, образуют разрыхленный хроматин, который распределён по всему объёму ядра. Разрыхление структуры хромосом – необходимое условие для транскрипции, передачи наследственной информации, содержащейся в ДНК. Разрыхленный хроматин может быть активным и неактивным в зависимости от процессов регуляции
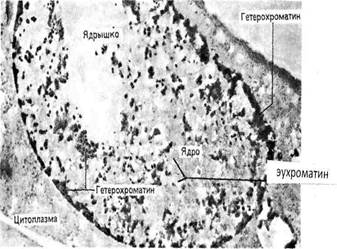 Классификация и функции хроматина: различают гетеро- и эухроматин.
Классификация и функции хроматина: различают гетеро- и эухроматин.
а) Гетерохроматин.Во время интерфазы определённые участки хромосом и целые хромосомы остаются компактными. Они образуют «глыбки» интенсивно окрашенные и, как правило, прилежащие к мембране ядра. Гетерохроматин неактивен в отношении транскрипции. Существует две формы гетерохроматина: факультативный и конститутивный
· факультативный гетерохроматин бывает гетерохроматичным только временами. Он информативен, содержит гены с которых считывается наследственная информация, когда гетерохроматин переходит в эухроматическое состояние. Образуется при спирализации одной из двух гомологичных хромосом. Типичным примером служит тельце полового хроматина, образуемого одной из двух Х-хромосом соматических клеток женских особей человека и млекопитающих. Функциональная роль факультативного гетерохроматина заключается в компенсации снижении дозы определенного гена (например, появление промежуточного признака при явлении неполного доминирования у гетерозигот Аа, влияет на экспрессивность проявления наследственных признаков в фенотип), определяет тканеспецифичность.
· структурный гетерохроматин – отличается высокоспирализованным состоянием, которое сохраняется на протяжении всего мит. цикла. Он занимает постоянные участки в гомологичных хромосомах – это фрагменты околоцентромерных, теломерных участков хромосом, Не содержит структурных генов (нетранскрибируемый); Его роль не ясна, но по видимому он выполняет опорную функцию.
б) Эухроматин (разрыхленный) - имеет менее компактную организацию, деспирализуется в конце митоза, образует слабоокрашенные нитчатые структуры содержит структурные транскрибируемые гены;
В КАЖДОЙ ХРОМОСОМЕ СВОЙ ПОРЯДОК РАСПОЛОЖЕНИЯ ЭУ- И ГЕТЕРОХРОМАТИНА, ЧТО ИСПОЛЬЗУЕТСЯ ДЛЯ ИДЕНТИФИКАЦИИ ОТДЕЛЬНЫХ ХРОМОСОМ В ЦИТОГЕНЕТИКЕ.
Уровни структурной организации хроматина:
Данные микроскопического и электронно-микроскопического изучения хроматина и митотических хромосом дают следующую картину структурной организации хромосом:
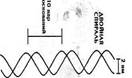
· двойная спираль ДНК - 1,5 нм (толщина биспирали)
· нуклеосомная нить (ДНК присоединяет белки и скручивается в нуклеогистоновый комплекс) - 8 молекул гистонов: Н2а, Н2в, Н3, Н4 они служат основой – образуя белковые тела - коры, на которые «накручены» фрагменты ДНК длиной примерно в 200 пар нуклеотидов. Гистон Н1 «сшивает» витки ДНК. Участки ДНК не связанные с белками, расположенные между гистоновыми корами, называются связующими или линкёрными. Результат скручивания ДНК и присоединение белка преобразуется в нуклеогистоновый комплекс с нуклеосомной структурой – 10 – 13 нм
· хроматиновая фибрилла 20 – 25 нм, дальнейшее скручивание ДНК и присоединение белков
· серии петельных доменов (хромонема) 100 – 200 нм. Домен – область с поперечным размером, возвышение
· конденсированный участок хромосомы – 700 нм (образуют глыбки хроматина)
· метафазная хромосома - 1400 нм
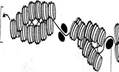
Линкёрная ДНК Гистоновый кор
Нуклеосомы
Дата добавления: 2019-12-09; просмотров: 923;
Поиск по сайту
Узнать еще
- A. Узагальнені координати і узагальнені швидкості та прискорення
- F-распределение Фишера–Снедекора
- II. 4. Скорость оседания эритроцитов (СОЭ)
- II. Корректура карт.
- II. Этап осознанной коррекции «новыми собственниками»
- III. Охлаждение с заданной скоростью.
- IV. Деятельность акционерного общества в интересах иных заинтересованных сторон и корпоративная социальная ответственность.
- IV.1.7. ПСИХОЛОГИЧЕСКАЯ ДИАГНОСТИКА И КОРРЕКЦИЯ ПРИ РАННЕМ ДЕТСКОМ АУТИЗМЕ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
