Эндокринный аппарат поджелудочной железы хордовых.
Поджелудочная железа - это орган смешанной секреции, поскольку содержит как экзокринную часть, секретирующую панкреотическй сок, так и эндокринный аппарат.
У ланцетника и круглоротых поджелудочная железа еще не имеет органного типа, но у них присутствуют железистые клетки, гомологичные экзо- и эндокриноцитам поджелудочной железы более высокоорганизованных животных. У ланцетника такого типа клетки располагаются между эпителиоцитами слизистой оболочки кишки и печеночного выроста (в дивертикуле), образуя эндоэпителиальную железу мозаично-клеточного типа (рис. 42). Эндокринные клетки неоднородны: одни из них синтезируют и выделяют гормоны глюкагон и гастрин, а другие – инсулин.
Вышеуказанные типы эндокринных клеток одним концом достигают просвета кишечника, а другим вступают в контакт с базальной мембраной, под которой располагаются кровеносные капилляры.
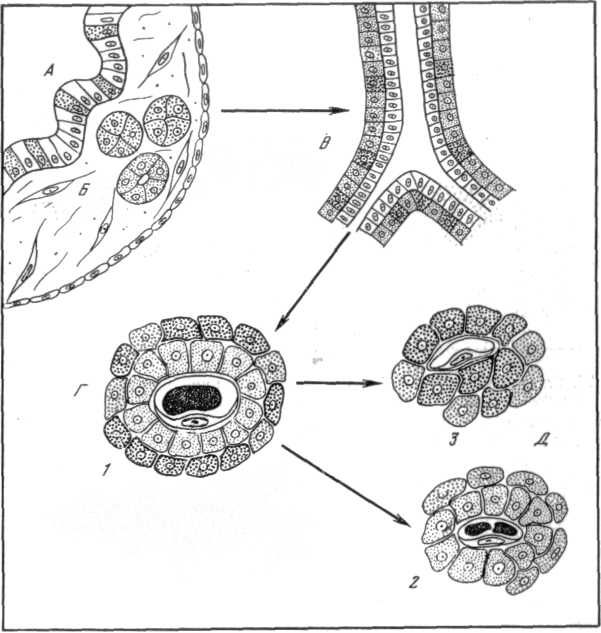
Рис.42. Схема этапов эволюции форм структурной организации эндокринной части поджелудочной железы позвоночных.
А – инсулярный аппарат бесчерепных (железа мозаично-клеточного типа); Б – инсулярный аппарат круглоротых (скопления и фолликулы Лангерганса); В – эндокринная часть поджелудочной железы хрящевых и кистеперых рыб («внешний эпителий»); Г – эндокринная часть поджелудочной железы костистых рыб, земноводных, пресмыкающихся, птиц и млекопитающих («смешанные» панкреатические островки); Д – эндокринная часть поджелудочной железы птиц («темные» и «светлые» панкреатические островки); 1 – А – клетка; 2 – В – клетка; 3 – Д –клетка
( по Яглову, 1978).
У круглоротых железистые клетки, гомологичные инсулярному аппарату поджелудочной железы более высокоорганизованных позвоночных, располагаются как в соединительнотканном слое самой кишки, так и в соединительной ткани, окружающей кишечник. Причем, в некоторых случаях описаны скопления клеток инсулярнного аппарата даже в печеночной паренхиме. Прямая связь эндокриноцитов с капиллярным руслом кишечника полностью завершается только у взрослых животных. Выделяют два основных скопления эндокринных клеток – краниальное (в начале передней кишки) и каудальнаое (в области впадения жёлчного протока).
Эмбриональным источником развития инсулярного аппарата у круглоротых является кишечный эпителий клетки, которого уже на ранних стадиях развития начинают мигрировать в собственный соединительнотканный слой слизистой оболочки. И хотя на данной стадии развития еще отсутствует прямая связь эндокринных клеток инсулярного аппарата с капиллярным руслом кишечника, еще в личиночном периоде начинается его функционирование. Окончательное формирование эндокринного аппарата завершается у взрослых особей после метаморфоза.
Одновременно с миграцией эндокринных клеток из кишечного эпителия идет процесс васкуляризации инсулярного аппарата, который полностью завершается только у взрослых особей. В центральных участках скоплений эндокринных клеток появляются заполненные коллоидоподобным веществом полости, и таким образом формируются фолликулы, получившие названия фолликулов Лангерганса.
Клеточный состав инсулярного аппарата неоднороден. В нем имеются гранулярные и агранулярные клетки. Основная масса гранулярных клеток представлена В-клетками, образующими инсулин. Роль других гранулярных клеток не ясна. Агранулярные клетки, видимо, осуществляют усвоение гормонов из коллоида фолликулов Лангерганса и обеспечивают их транспорт в капиллярное русло. Однако, это предположение остается недоказанным. Нет данных и о наличии А – клеток, образующих глюкагон.
Впервые поджелудочная железа приобретает органный статус у хрящевых рыб. У них это компактный орган дольчатого строения, состоящий из экзокринного и эндокринного отделов. Эндокринные клетки располагаются на ходу средних и мелких выводных протоков. У некоторых видов хрящевых рыб имеются другие формы организации эндокринной части железы: у одних эндокринные клетки окружают выводные протоки их в виде эпителиального слоя («внешний эпителий»); у других, кроме этого, имеются скопления эндокринных клеток в виде прилежащих к выводным протокам столбов; у третьих – помимо эндокринных клеток, располагающихся вокруг протоков, встречаются изолированные островки таких клеток.
У хрящевых рыб в отличие от круглоротых впервые появляются три вида самостоятельных гормонопродуцирующих клеток -А, В, и Д. У подкласса цельноголовых описан еще и четвертый тип клеток, названный Х – клетками. Гормональный профиль идентифицирован только для А - , В- и Д - клеток. Клетки А продуцируют глюкагон, В-инсулин, а Д-соматостатин. Последний оказывает ингибирующий эффект на экзокринный отдел железы и на выделение гормонов А - и В-клетками.
У костных рыб поджелудочная железа имеет характер компактного органа только у хищных, а у остальных рыб она представлена гроздьями железистой ткани, разбросанными в брыжейке и других органах.
У латимерии имеются скопления эндокринных клеток как по ходу средних и мелких выводных протоков, а также в виде свободно расположенных островков, не связанных с выводными протоками. Эндокринный отдел представлен А- , В- и Д-клетками.
У двоякодышащих рыб эндокринная часть поджелудочной железы имеет вид только островков А- и В- эндокриноцитов. Такой же тип организации эндокринного отдела характерен и для костистых рыб. Клеточный состав эндокринных островков у них изучен недостаточно: в них идентифицировано от одного до шести видов секреторных клеток. У большинства изученных видов идентифицированы А -, В - и Д- клетки.
Таким образом, бесчерепные были первичными хордовыми, у которых появился инсулярный аппарат, представленный В-клетками, располагающимися в эпителии слизистой оболочки кишечника. Помимо В-клеток, здесь присутствуют клетки, секретирующие энтероглюкагон и гастрин. У круглоротых произошло выселение популяции В-клеток в собственно соединительнотканный слой слизистой оболочки или даже за пределы кишки. При этом изменился и характер структурной организации инсулярного аппарата, который помимо скоплений эндокринных клеток стал содержать и фолликулы Лангерганса.
У хрящевых рыб сформировался компактный орган, состоящий из экзокринного и эндокринного отделов. Последний пополнился новыми гормонопродуцирующими клетками (А -, Д -, а у некоторых и Х-клетками). Если В-клетки специализированы на выработку инсулина, то А-клетки – на контринсулярном гормоне – глюкагоне. Д-клетки секретируют фактор местной регуляции эндокринного аппарата поджелудочной железы – соматостостатин.
У хрящевых и кистеперых рыб основная масса эндокриноцитов сосредоточена по ходу выводных протоков, что явилось переходом к следующей эволюционной форме – панкреатическим островкам.
Земноводные -следующий этапом в эволюции позвоночных. Они произошли от кистеперых рыб и являются первыми примитивными наземными животными. Морфология эндокринного отдела поджелудочной железы у них изучена еще недостаточно полно, большинство исследований выполнены на представителях отряда бесхвостых. Эндокринная часть поджелудочной железы у всех изученных видов амфибий организована в виде островков. Многими исследователями в панкреатических островках были обнаружены А -, В - и Д - клетки, а у некоторых видов из отряда хвостатых описаны еще два дополнительных типа клеток. Обращает на себя внимание общность структурной организации панкреатических островков у разных видов амфибий: в центре островка находится синусоидальный капилляр, вокруг которого располагаются В-клетки, тогда как А- и Д-клетки лежат по его периферии.
Дляпресмыкающиесяхарактерен сходный с амфибиями принцип организации островков эндокриноцитов. У них островки располагаются в толще железы. В то же время у змей такого рода островки обнаруживаются еще и в селезенке. У некоторых видов черепах, помимо панкреатических островков, имеются тяжи А - клеток, располагающихся по ходу мелких выводных протоков. В островках идентифицированы А -, В - и Д - клетки. Однако, есть сведения о наличии четвертого типа клеток, гормональный, профиль которых еще не выяснен.
В поджелудочной железептицобнаружены эволюционно новые формы организации эндокринного аппарата. У них имеются так называемые «темные» островки эндокриноцитов, состоящие из А - клеток и только единичных В - клеток. Кроме этого, есть панкреатические островки, состоящие из В-клеток и только единичных Д-клеток. Они называются «светлыми» островками. Таким образом, у птиц существует разделение инсулярного и контринсулярного компонентов эндокринного аппарата. По-видимому, это вызвано необходимостью поддержания высокого уровня глюкозы в крови в связи с интенсивными процессами метаболизма. Однако, у некоторых видов птиц( например, у куриц и голубя) эндокринная часть железы характеризуется наличием «темных», «светлых» и «смешанных» островков. «Смешанные» состоят из А-, В- и Д-клеток. Видимо, эволюционно более древние «смешанные» островки были дополнены эволюционно более поздними «темными» и «светлыми». Это может рассматриваться как приспособительная перестройка эндокринной части поджелудочной железы к экологическим особенностям птиц, требующим высокого содержания в крови глюкозы в связи с высокой интенсивностью у птиц метаболических процессов.
У всех млекопитающих эндокринная часть поджелудочной железы организована в виде панкреатических островков (островки Лангерганса). Суммарно клеточный состав этого отдела железы у различных видов млекопитающих представлен: А - клетками (или по другой терминологии ά - клетками), В (или β) - клетками, Д (или G) – клетками, РР – клетками и δ – клетки). Но только А - , В – и Д – клетки обнаружены у всех млекопитающих. При этом А – клетки у большинства видов млекопитающих составляют 24 – 27% популяции клеток панкреатических островков, а у человека 15%. Они продуцируют глюкагон, котрый депонируется в их секреторных гранулах, проявляющих оксифильные свойства. Глюкагон стимулирует расщепление гликогена и жиров, что ведет к быстрой мобилизации энергии.
В (или β) - клетки составляют около 70% и располагаются преимущественно в центре островков. Их гранулы проявляют базофильные свойства. Они продуцируют инсулин, который, воздействуя на рецепторы клеток – мишеней (это в основном печень, скелетные мышцы, жировые клетки), является главным регулятором гомеостаза глюкозы. Он стимулирует мембранный транспорт глюкозы и образование гликогена, а также участвует в регуляции липидного и белкового обмена. При уменьшении синтеза инсулина или нарушении в клетках - мишенях рецепторов инсулина развивается сахарный диабет.
Д (или G) – клетки секретируют гастрин, стимулирующий секрецию соляной кислоты в желудке. Некоторые опухоли поджелудочной железы приводят к резкому увеличению уровня гастрина.
РР-клетки (F – клетки согласно другой терминологии) секретируют панкреатический полипептид – один из регуляторов пищевого режима за счет угнетения секреции панкреатического сока.
δ – клетки островков Лангерганса секретируют гормон соматостатин, который в поджелудочной железе подавляет гормональную активность образования инсулина и глюкагона.
Следует заметить, что соматостатин синтезируется и многими нейронами центральной нервной системы, а также клетками ряда внутренних органов. Он является мощным регулятором функций эндокринной и нервной систем. Так, например, ингибирует синтез и секрецию гормона роста, адренокортикотропного гормона и тиротропина в гипофизе.
Дата добавления: 2019-12-09; просмотров: 1199;
Поиск по сайту
Узнать еще
- IV. Движение поездов при неисправности электрожезловой системы и порядок регулировки количества жезлов в жезловых аппаратах
- А.7 Устройство и принципы действия адсорбционных аппаратов
- Абсцесс предстательной железы
- Автоматика газовых аппаратов
- Аденома предстательной железы
- Активные потери энергии в электрических аппаратах
- Акустически-резонаторная система голосового аппарата. Занятия с детьми в мутационный период
- Алгоритм технологического расчета теплообменных аппаратов

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
