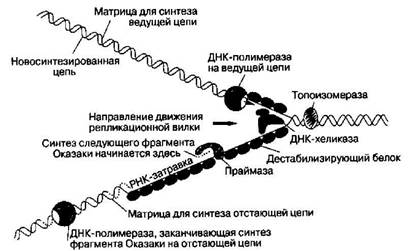
Расположение основных белков в репликационной вилке
|
Молекула ДНК-праймазы непосредственно связана с ДНК-хеликазой, образуя структуру, называемую праймосомой. Праймосома движется в направлении раскрывания репликационной вилки и по ходу движения синтезирует РНК-затравку для фрагментов Оказаки. В этом же направлении движется ДНК-полимераза ведущей цепи и ДНК-полимераза отстающей цепи. Для этого, как полагают, последняя накладывает цепь ДНК, которая служит ей матрицей, саму на себя, что и обеспечивает разворот ДНК-полимеразы отстающей цепи на 180 градусов. Согласованное движение двух молекул ДНК-полимераз обеспечивает координированную репликацию обеих нитей.
Всего в репликационной вилке одновременно работает около двадцати разных белков.
Процесс репликации хромосомы бактерий начинается в точке начала репликации и продолжается до тех пор, пока не удвоится вся ДНК хромосомы.
Молекулярно-биологические процессы, происходящие во время
репликации ДНК, в основном похожи у эукариот и прокариот. Если
бактериальная хромосома представляет собой единицу репликации – репликон,
то репликация ДНК эукариотической хромосомы осуществляется посредством разделения ее на множество отдельных репликонов. Полагают, что у эукариот гомологами ориджинов начала репликации являются автономно реплицирующиеся последовательности или ARS (autonomously replicating sequences).
По эукариотической хромосоме в каждый момент времени может двигаться независимо друг от друга множество репликационных вилок.Остановка продвижения вилки происходит только при столкновении с другой вилкой, движущейся во встречном направлении, или по достижении конца хромосомы. В результате вся ДНК хромосомы в короткий срок оказывается реплицированной.
Репликоны у эукариот имеют существенно меньшие размеры, чем у прокариот (хотя в пределах генома одного вида они могут варьировать в размерах в 10 раз). Скорость репликации существенно ниже у эукариот.
Репарация ДНК
В любой клетке под влиянием различных факторов в ДНК ежедневно происходят тысячи случайных изменений, а за год в каждой клетке накапливается лишь очень небольшое число стабильных изменений нуклеотидной последовательности ДНК. Среди множественных случайных замен оснований в ДНК лишь одна на тысячу приводит к возникновению мутации. Все остальные повреждения очень эффективно ликвидируются в процессе репарации ДНК. Механизм репарации («залечивание» повреждений ДНК) основан на том, что молекула ДНК имеет две копии генетической информации – по одной в каждой из нитей молекулы. Основной путь репарации включает три этапа:
- измененный участок поврежденной цепи ДНК распознается и удаляется с помощью ДНК-репарирующих нуклеаз. В спирали ДНК в этом месте возникает брешь;
- ДНК-полимераза и гликозилазы заполняют эту брешь, присоединяя нуклеотиды один за другим, копируя информацию с целостной нити;
- ДНК-лигаза «сшивает» разрывы и завершает восстановление молекулы.
Нарушение репарации у людей, пораженных пигментной ксеродермой (повышенная чувствительность к солнечному свету), приводит к раку кожи. Активность УФ-эндонуклеазы, необходимая для репарации, при этом отсутствует. Если подавляются репарационные системы, то мутагенез усиливается.
Литература
1. Айала, Ф. Современная генетика / Ф. Айала, Дж. Кайгер. – М.: Мир, 1987. – Т.1. – 295 с; Т.2. – 368 с; Т.3.
2. Дубинин, Н. П. Общая генетика / Н. П. Дубинин. – М.: Наука, 1986.
3. Жимулев, И. Ф. Общая и молекулярная генетика / И. Ф. Жимулев. – Новосибирск: Изд-во Новосибирского ун-та, 2002.
Транскрипция
Передача информации от ДНК осуществляется посредством информационной или матричной рибонуклеиновой кислоты (мРНК). Синтез мРНК называется транскрипцией. Молекула мРНК, комплементарная одной из цепей матричной ДНК, образуется в ходе сополимеризации четырех рибонуклеозидтрифосфатов (аденин-, гуанозин-, цитозин- и урацилтрифосфата) с образованием 3¢-5¢-фосфодиэфирных связей и освобождением неорганического пирофосфата.Транскрипцию осуществляет фермент ДНК-зависимая РНК-полимераза. Синтез мРНК молекулами РНК-полимеразы начинается в определенных местах ДНК, называемых промоторами, и завершается на особых нуклеотидных последовательностях – терминаторах. Совокупность нуклеотидов ДНК, заключенных между промотором и терминатором, называют транскрипционной единицей или транскриптоном.
Процесс транскрипции подразделяют на 4 стадии: связывание РНК-полимеразы с ДНК и распознавание промотора, инициация, элонгация и терминация. Предполагается, что после первоначального непрочного связывания с ДНК в случайном месте молекула РНК-полимеразы перемещается вдоль двойной спирали ДНК до тех пор, пока не обнаружит последовательность нуклеотидов промотора. В этом месте связывание молекулы фермента с ДНК становится более прочным.Инициация транскрипции начинается с образования на промоторе предъиниционного комплекса, состоящего из РНК-полимеразы и матричной ДНК. После сборки предъинициационный комплекс претерпевает температурно-зависимые конформационные изменения, которые сопровождаются локальным плавлением, то есть расплетением двойной спирали ДНК, и комплекс становится способным к транскрипции. При наличии рибонуклеозидтрифосфатов происходит образование первых фосфодиэфирных связей в молекуле синтезируемой мРНК, после чего начинается стадия элонгации, то есть последовательное удлинение синтезируемой молекулы мРНК.
В 1992 г. М. Чэмберлен с сотрудниками разработали общую модель элонгации мРНК, согласно которой перемещение РНК-полимеразы вдоль ДНК и присоединение нуклеотидов к растущей цепи мРНК в активном центре фермента разделены во времени. Это разделение возможно потому, что у РНК-полимеразы имеется два сайта (участка), удерживающих растущую цепь мРНК, и два участка связывания ДНК-матрицы. Молекула РНК-полимеразы перемещается вдоль ДНК подобно гусенице: когда один сайт связывания ДНК фиксирован, другой перемещается вперед. ДНК-зависимые РНК-полимеразы фагов, состоящие из одной субъединицы, синтезируют РНК в условиях in vitro со скоростью 200-400 нуклеотидов в секунду. При перемещении фермента вдоль матрицы цепи ДНК подвергаются плавлению и повторному отжигу, в результате которого восстанавливается исходная структура ДНК. Стадия элонгации заканчивается после достижения РНК-полимеразой терминатора транскрипции.Затем синтезированная РНК и РНК-полимераза освобождаются из транскрипционного комплекса. Только минус-цепь ДНК служит матрицей для синтеза мРНК.
Участки ДНК, несущие информацию о строении белка – экзоны, разделены неинформативными интронами. В процессе транскрипции считывается информация как с экзонов, так и с интронов. Образуется предшественник мРНК - про-мРНК. Молекулы про-мРНК претерпевают созревание – процессинг. В ядре из про-мРНК происходит вырезание интронов и объединение экзонов – сплайсинг.После этого мРНК соединяется с белком, образуя инфорсому. Она выходит через поры в ядерной оболочке в цитоплазму. мРНК высвобождается из инфорсомы и одноцепочечная неспирализованная молекула мРНК присоединяется к участку малой субъединицы рибосомы, который примыкает к большой субъединице. К рибосоме прикрепляется небольшой участок цепи мРНК, содержащий один кодон, состоящий из трех азотистых оснований. Один кодон соответствует одной аминокислоте.
Генетический код
В любом данном участке ДНК только одна из двух нитей ДНК кодирует аминокислоты, поэтому код – это последовательность нуклеотидов, а не пар нуклеотидов. Генетический код имеет следующие свойства:
1. Генетический код читается группами по три нуклеотида, т.е. код триплетный. Каждый триплет кодирует аминокислоту, каждый триплет называется кодоном.
2. Основные закономерности организации генетического кода были открыты с помощью генетического анализа района II фага Т6. В 1961 г. Ф. Крик и его коллеги показали, что код должен читаться неперекрывающимися триплетами с фиксированной стартовой точки.
а) неперекрывание подразумевает, что каждый кодон состоит из трех нуклеотидов и каждый последующий кодон представлен следующими тремя нуклеотидами.
б) Фиксированная стартовая точка означает, что считывание начинается на одном конце и завершается на другом; различные части кодирующей последовательности не могут считываться независимо друг от друга.
Началом трансляции любого гена является кодон AUG. В конце гена обязательно стоят кодоны UAA, UAG или UGA, которые не кодируют аминокислот и являются сигналами на окончание синтеза белка - стоп-кодоны. Для повышения надежности процесса терминации стоп-кодоны обычно дублируются. Первым при этом, как правило, выступает кодон UAA (основной терминирующий триплет), а вслед за ним на очень
близком расстоянии в той же рамке считывания следует один из запасных
терминирующих триплетов - UAG или UGA.
Таблица –Соответствие кодонов генетического кода аминокислотам белка
| Первая буква в кодоне (5') | Вторая буква в кодоне | Третья буква в кодоне (3') | |||
| U | С | А | G | ||
| U | Фен (F) Фен (F) Лей (L) Лей (L) | Сер (S) Сер (S) Сер (S) Сер (S) | Тир (Y) Тир (Y) Stop Stop | Цис (С) Цис (С) Stop Трп (W) | U с А G |
| С | Лей (L) Лей (L) Лей (L) Лей (L) | Про (Р) Про (Р) Про (Р) Про (Р) | Гис (Н) Гис (Н) Глн (Q) Глн (Q) | Apr (R) Apr (R) Apr (R) Apr (R) | U С А G |
| А | Иле (I) Иле (I) Иле (I) Мет (М) | Тре (Т) Тре (Т) Тре (Т) Тре (Т) | Ach(N) Ach(N) Лиз (К) Лиз (К) | Сер (S) Сер (S) Apr (R) Apr (R) | U С А G |
| G | Вал(У) Вал(У) Вал(У) Вал(У) | Ала (А) Ала (А) Ала (А) Ала (А) | Асп (D) Асп (D) Глу (Е) Глу (Е) | Гли (G) Гли (G) Гли (G) Гли (G) | U С А G |
Примечание
U - урацил, С - цитозин, А - Аденин, G - гуанин, F, L, I и т.д. - однобуквенные сокращения названий аминокислот, Фен, Сер и т.д. -трехбуквенные сокращения. Кодоны UAA, UAG, UGA - не кодируют аминокислот. Эти кодоны являются сигналами терминации трансляции - стоп-кодонами
в) Если генетический код считывается неперекрывающимися триплетами, есть только три возможности транслирования нуклеотидной последовательности в аминокислотную, в зависимости от стартовой точки. Эти три возможности называют рамками считывания.
Мутация, в результате которой инсертируется или делетируется один нуклеотид и изменяется рамка считывания всей последующей последовательности, называется сдвигом рамки. Поскольку последовательность новой рамки считывания полностью отлична от первоначальной, вся аминокислотная последовательность будет измененной ниже мутации. Функция такого белка полностью утрачена.
Дополнение
Проблема кодирования в молекулярной биологии была впервые поставлена Г. Гамовым еще в начале 50х годов, т.е. задолго до открытия самой мРНК. Размышляя над тем, как линейная последовательность четырех различных нуклеотидов может определить последовательность двадцати разных аминокислот в белке, Гамов предположил, что генетический код является триплетным. Он же поставил вопросы и о других свойствах генетического кода: перекрываемости, запятых между кодонами, вырожденности.
3. Генетический код является вырожденным, в том смысле, что одной аминокислоте может соответствовать несколько кодонов (Таблица). Однако, кодоны используются не с одинаковой частотой. Например, у дрозофилы, в результате параллельного изучения последовательностей кодонов в генах и аминокислот, кодированных ими, при этом суммарная длина генов соответствовала 269 т.п.н., было показано, что кодоны используются с разными частотами.
Дополнение
В 1968 году за открытие и интерпретацию генетического кода и его функции в белковом синтезе Нобелевская премия была присуждена Р. Холли, X. Г. Хоране и М.В. Ниренбергу.
4. Генетический код универсален, в том смысле, что определённому кодону соответствует определённая аминокислота. Например, AUG-кодон кодирует метионин у любого организма. Однако, по мере расширения круга объектов молекулярной генетики стали накапливаться исключения, сделавшие код "квазиуниверсальным". Касается это прежде всего митохондриальных геномов.
Трансляция
Трансляция (перевод) – процесс воплощения генетической информации мРНК в структуру полипептида. Это второй этап белкового синтеза, осуществляемый последовательной поликонденсацией отдельных аминокислотных остатков, начиная с аминоконца полипептидной цепи к карбоксильному концу.
Зрелая матричная РНК выходит в цитоплазму, где осуществляется процесс транскрипции – декадирование мРНК в аминокислотную последовательность белка. Процесс декадирования осуществляется в направлении от 5`→3` и происходит в рибосомах. Комплекс мРНК и рибосом называется полисомой. Подобно транскрипции механизм трансляции состоит из трех этапов: инициации, элонгации и терминации.
Трансляция начинается со стартового кодона АУГ, который при локализации в смысловой части структурного гена кодирует аминокислоту метионин. Каждую аминокислоту доставляет к полисоме транспортная РНК (тРНК), специфичная к данной аминокислоте. тРНК выполняет роль посредника между кодоном мРНК и аминокислотой. Молекулы тРНК узнают в цитоплазме соответствующий триплет (кодон в мРНК) по принципу спаривания комплементарных азотистых оснований. тРНК, которая подходит к малой субчастице, образует связь кодон-антикодон, при этом одновременно передает свою аминокислоту в аминоацильный участок (А-участок) большой субъединице. К кодону АУГ «подходит» антикодон только той тРНК, которая переносит метионин. Поэтому прежде всего к рибосоме доставляется метионин. Затем кодон АУГ переходит на пептидильный участок большой субъединицы (Р-участок). В результате этих процессов образуется транслирующая рибосома – инициирующий комплекс.
Элонгация – это последовательное включение аминокислотных остатков в состав растущей полипептидной цепи. Каждый акт элонгации состоит из трех этапов:
- узнавание кодона, которое заключается в связывании антикодона с очередной молекулой тРНК, несущей аминокислоту, с кодоном свободного А-участка на рибосоме;
- образование пептидной связи, которое происходит лишь тогда, когда оба участка А и Р заняты молекулами тРНК. Часть большой субъединицы рибосомы – фермент пептидилтрансферазу, катализирующий образование пептидной связи;
- транслокация, где тРНК участка Р, не связанная с пептидом, покидает рибосому. Затем молекула тРНК с полипептидом переходят из А на Р-участок и, наконец, рибосома перемещается вдоль РНК на один кодон.
Терминация (окончание синтеза) происходит по команде кодонов УАА, УАГ, УГА. В природе не существует таких молекул тРНК, антикодоны которых соответствовали бы этим кодонам.
Каждая мРНК транслируется, как правило, несколько раз, после чего разрушается. Среднее время жизни молекулы мРНК около 2 мин. Разрушая старые и образуя новые мРНК, клетка может довольно строго регулировать как тип продуцируемых белков, так и их количество. Это регуляция синтеза белка на уровне транскрипции. У эукариот возможна регуляция и на уровне трансляции.
Синтез белка – один из существеннейших показателей жизни.
Дата добавления: 2017-03-12; просмотров: 2386;
Поиск по сайту
Узнать еще
- А) позиция непринужденного общения (угловое расположение).
- Автомобильные генераторы – методика поиска основных
- АМИНОКИСЛОТНЫЕ СМЕСИ И БЕЛКОВЫЕ ГИДРОЛИЗАТЫ
- Аминокислотный состав белков
- АМИНОКИСЛОТНЫЙ СОСТАВ БЕЛКОВ
- Амортизация основных производственных фондов
- Амортизация основных средств.
- Амортизация основных фондов предприятия

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
