развитие теории Хейфлика
Теории старения
Мысль о том, что старение может быть заложено с момента рождения, была высказана немецким ученым-дарвинистом Августом Вейсманом (Friedrich Leopold August Weismann, 1834-1914). В своей знаменитой лекции, прочитанной в 1891 году, Вейcман выдвинул предположение, что смерть от старости возникла в ходе эволюции: "Я рассматриваю смерть не как первичную необходимость, а как нечто приобретенное вторично в процессе адаптации."
ПОДХОДЫ К КЛАССИФИКАЦИИ ТЕОРИЙ СТАРЕНИЯ
Теории, объясняющие старение организмов можно классифицировать различными способами.
Например, существует разделение на три группы: генетические теории, в которых генно-контролируемые запрограммированные <биологические часы>, такие как теломеры регулируют рост, зрелость и старость, нейроэндокринные теории и теории накопления повреждений. Вообще говоря, это разделение довольно условное, потому как все эти механизмы важны и взаимосвязаны.
Также выделяют 2 большие группы: стохастические (вероятностные) теории и теории программированного старения.
Можно классифицировать теории по уровню организации живой материи.
По мнению В.Н. Анисимова, руководителя Российского Геронтологического Общества, наиболее яркими теориями остаются выдвинутая в 1956 г. Д. Харманом свободнорадикальная теория (Harman, 1956, 1998), теория клеточного (репликативного) старения Л. Хейфлика (Hayflick, Moorhead, 1961; Hayflick, 1998), теломерная теория А.М.Оловникова (Оловников, 1971; Olovnikov, 1996), элевационная теория старения В.М. Дильмана (Дильман, 1987; Dilman, 1971, 1994) и теория расходуемой сомы Т. Кирквуда (Kirkwood, 1997, 2002). выдвинутая в 1956 г. Д. Харманом свободнорадикальная теория, теория клеточного (репликативного) старения Л. Хейфлика и теломерная теория А. М. Оловникова, элевационная теория старения В. М. Дильмана.
КЛАССИФИКАЦИЯ ТЕОРИЙ СТОХАСТИЧЕСКОГО СТАРЕНИЯ
(Schulz-Aellen, 1997)
· Теория cоматических мутаций - Соматические мутации нарушают генетическую информацию и уменьшают функцию клеток
· Катастрофа ошибок - Ошибки процессов транскрипции и/или трансляции уменьшают эффективность клеток
· Повреждения ДНК, репарация ДНК - Повреждения ДНК постоянно репарируются различными механизмами. Эффективность репарации положительно корелирует с продолжительностью жизни и уменьшается с возрастом
· Повреждения белков - Конформационные нарушения белков и ферментов (перекрестные сшивки) повреждают функцию клетки
· Перекрестные сшивки - Химические перекрестные сшивки важных макромолекул (например, коллагена) приводят к нарушениям функции клеток и тканей
· Износ - Накопление повреждений в повседневной жизни уменьшает эффективность организма
КЛАССИФИКАЦИЯ ТЕОРИЙ ПРОГРАММИРОВАННОГО СТАРЕНИЯ
(Schulz-Aellen, 1997)
· Генетические теории - Старение вызывается запрограммированными изменениями экспрессии генов, или экспрессией специфических белков
· Гены смерти - Существуют гены клеточной гибели
· Избирательная гибель - Гибель клетки обусловлена наличием специфических мембранных рецепторов
· Укорочение теломер - Укорочение теломер с возрастом in vitro и in vivo приводит к нестабильности хромосом и гибели клеток
· Нарушения дифференцировки - Ошибки в механимзах активации-репрессии генов, приводящие к синтезу избыточных, несущественных или ненужных белков
· Накопление <загрязнений> - Накопление отходов метаболизма снижает жизнеспособность клеток
· Нейроэндокринные теории - Недостаточность нервной и эндокринной систем в поддержании гомеостаза. Потеря гомеостаза приводит к старению и смерти
· Иммунологическая теория - Определенные аллели могут увеличивать или сокращать продолжительность жизни.
· Метаболические теории - Долголетие обратно пропорционально скорости метаболизма
· Свободно-радикальная теория - Долголетие обратно пропорционально степени повреждения свободными радикалами и прямо пропорционально эффективности антиокислительных систем
· Часы старения - Старение и смерть являются результатом предопределенного биологического плана
· Эволюционные теории - Естественный отбор устраняет индивидуумов после того, как они произведут потомство
КЛАССИФИКАЦИЯ ВАЖНЕЙШИХ ТЕОРИЙ СТАРЕНИЯ ПО УРОВНЮ ИНТЕГРАЦИИ
(Yin, Chen, 2005)
Организменный уровень интеграции
Теория изнашивания - Sacher, 1966
Теория катастрофы ошибок - Orgel, 1963
Теория стрессового повреждения - Stlye, 1970
Теория аутоинтоксикации - Metchnikoff, 1904
Эволюционная теория (теория программированного старения) - Williams, 1957
Теория сохранения информации (теория программированного старения)
Органный уровень
Эндокринная теория - Korenchevsky, 1961
Иммунологическая теория - Walford, 1969
Торможение головного мозга
Клеточный уровень
Теория клеточных мембран - Zg-Nagy, 1978
Теория соматических мутаций - Szillard, 1959
Митохондриальная теория - Miquel et al., 1980
Митохондриально-лизосомальная теория - Brunk, Terman, 2002
Теория пролиферативного лимита клетки (теория программированного старения) - Hayflick, Moorhead, 1961
Молекулярный уровень
Теория накопление повреждений ДНК - Vilenchik, 1970
Теория следовых элементов - Eichhorn, 1979
Свободно-радикальная теория - Harman, 1956
Теория поперченных сшивок - Bjorksten, 1968
Теория окислительного стресса - Sohal, Allen, 1990; Yu, Yang, 1996
Теория неэнзиматической гликозиляции - Cerami, 1985
Теория карбонильной интоксикации - Yin, Brunk, 1995
Теория катастрофы загрязнения - Terman, 2001
Теория генных мутаций
Теория укорочения теломер (теория программированного старения) - Оловников, 1971
Прочие подходы
Cтарение как энтропия - Sacher, 1967; Bortz, 1986
Математические теории и различные унифицированные теории - Sohal, Alle, 1990;
Zg-Nagy, 1991; Kowald, Kirkwood, 1994
СВОБОДНОРАДИКАЛЬНАЯ ТЕОРИЯ СТАРЕНИЯ ДЭНХЕМА ХАРМАНА

Свободнорадикальная теория старения была представлена в 1956 Денхамом Харманом, который предположил, что старение является результатом случайного вредоносного повреждения тканей свободными радикалами. В ходе жизнедеятельности каждой клетки через неё проходит огромное количество кислорода. Он используется для клеточного дыхания, дающего клетке энергию. Но небольшая доля кислорода при этом уходит в паразитные соединения, обладающие огромной реакционной способностью. Их называют АФК - активные формы кислорода (хотя в их составе бывает и не только кислород). Примерами таких веществ могут служить всем хорошо известные перекись водорода и озон. Эти два вещества, однако, относительно малоактивны и могут существовать долго. Другие же АФК несравненно агрессивней. В организме они живут лишь тысячные доли секунды. А потом вступают в реакцию с другими молекулами, повреждая их. Они атакуют белки, липиды клеточных мембран, ДНК...
В результате атак со стороны АФК повреждаются митохондрии. Накопление этих повреждений и является сутью старения.
ТЕОРИЯ КЛЕТОЧНОГО СТАРЕНИЯ ЛЕОНАРДА ХЕЙФЛИКА

В 1961 г. Леонард Хейфлик представил данные о том, что даже в идеальных условиях культивирования фибробласты эмбриона человека способны делиться только ограниченное число раз (50 + 10). Последняя фаза жизни клеток в культуре была уподоблена клеточному старению, а сам феномен получил по имени автора название "предела Хейфлика". Сам Хейфлик не предложил объяснение этого явления.
ТЕЛОМЕРНАЯ ТЕОРИЯ ОЛОВНИКОВА
развитие теории Хейфлика

В 1971 г. научный сотрудник Института биохимической физики РАН А.М. Оловников, используя данные о принципах синтеза ДНК в клетках, предложил гипотезу, по которой <предел Хейфлика> объясняется тем, что при каждом клеточном делении хромосомы немного укорачиваются. У хромосом имеются особые концевые участки - теломеры, которые после каждого удвоения хромосом становятся немного короче, и в какой-то момент укорачиваются настолько, что клетка уже не может делиться. Тогда она постепенно теряет жизнеспособность - именно в этом, согласно теломерной теории, и состоит старение клеток. Открытие в 1985 г. фермента теломеразы, достраивающего укороченные теломеры в половых клетках и клетках опухолей, обеспечивая их бессмертие, стало блестящим подтверждением теории Оловникова. Правда, предел в 50-60 делений справедлив далеко не для всех клеток: раковые и стволовые клетки теоретически могут делиться бесконечно долго, в живом организме стволовые клетки могут делиться не десятки, а тысячи раз, но связь старения клеток с укорочением теломер является общепризнанной.
ЭЛЕВАЦИОННАЯ ТЕОРИЯ СТАРЕНИЯ
Выдвинута и обоснована в начале 50-х годов прошлого века ленинградским ученым Владимиром Дильманом. Согласно этой теории, механизм старения начинает свою работу с постоянного возрастания порога чувствительности гипоталамуса к уровню гормонов в крови. В итоге увеличивается концентрация циркулирующих гормонов. Как результат, возникают различные формы патологических состояний, в том числе характерные для старческого возраста: ожирение, диабет, атеросклероз, канкриофилия, депрессия, метаболическая имуннодепрессия, гипертония, гиперадаптоз, автоиммунные заболевания и климакс. Эти болезни ведут к старению и в конечном итоге к смерти.
Другими словами, в организме, существуют большие биологические часы, которые отсчитают отпущенное ему время жизни от рождения до смерти. Эти часы в определенный момент запускают деструктивные процессы в организме, которые принято называть старением.
По Дильману, старение и связанные с ним болезни - это побочный продукт реализации генетической программы онтогенеза - развития организма.
Из онтогенетической модели следует, что если стабилизировать состояние гомеостаза на уровне, достигаемом к окончанию развития организма, то можно затормозить развитие болезней и естественных старческих изменений и увеличить видовые пределы жизни человека.
ТЕОРИЯ РАСХОДУЕМОЙ (ОДНОРАЗОВОЙ) СОМЫ
ТЕОРИЯ ПЕРЕКРЕСТНЫХ СШИВОК
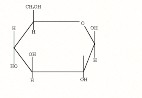
Этот механизм старения немного похож на воздействие свободных радикалов. Только роль агрессивных веществ здесь играют сахара, в первую очередь - всегда присутствующая в организме глюкоза. Сахара могут вступать в химическую реакцию с различными белками. При этом, естественно, функции этих белков могут нарушаться. Но что гораздо хуже, молекулы сахаров, соединяясь с белками, обладают способностью <сшивать> молекулы белков между собой. Из-за этого клетки начинают хуже работать. В них накапливается клеточный мусор.
Одно из проявлений такой сшивки белков - потеря тканями эластичности. Внешне наиболее заметным оказывается появление на коже морщин. Но гораздо больший вред приносит потеря эластичности кровеносных сосудов и лёгких. В принципе, у клеток есть механизмы для разрушения подобных сшивок. Но этот процесс требует от организма очень больших энергозатрат.
Сегодня уже существуют лекарственные препараты, которые разбивают внутренние сшивки и превращают их в питательные вещества для клетки.
ТЕОРИЯ ОШИБОК

Гипотеза <старения по ошибке> была выдвинута в 1954 г. американским физиком М. Сциллардом. Исследуя эффекты воздействия радиации на живые организмы, он показал, что действие ионизирующего излучения существенно сокращает срок жизни людей и животных. Под воздействием радиации происходят многочисленные мутации в молекуле ДНК и инициируются некоторые симптомы старения, такие как седина или раковые опухоли. Из своих наблюдений Сцилард сделал вывод, что мутации являются непосредственной причиной старения живых организмов. Однако он не объяснил факта старения людей и животных, не подвергавшихся облучению.
Его последователь Л. Оргель считал, что мутации в генетическом аппарате клетки могут быть либо спонтанными, либо возникать в ответ на воздействие агрессивных факторов - ионизирующей радиации, ультрафиолета, воздействия вирусов и токсических (мутагенных) веществ и т.д. С течением времени система репарации ДНК изнашивается, в результате чего происходит старение организма.
ТЕОРИЯ АПОПТОЗА (САМОУБИЙСТВА КЛЕТОК)

Академик В.П. Скулачев называет свою теорию теорией клеточного апоптоза. Апоптоз (греч. <листопад>) - процесс запрограммированной гибели клетки. Как деревья избавляются от частей, чтобы сохранить целое, так и каждая отдельная клетка, пройдя свой жизненный цикл, должна отмереть и ее место должна занять новая. Если клетка заразится вирусом, или в ней произойдет мутация, ведущая к озлокачествлению, или просто истечет срок ее существования, то, чтобы не подвергать опасности весь организм, она должна умереть. В отличие от некроза - насильственной гибели клеток из-за травмы, ожога, отравления, недостатка кислорода в результате закупоривания кровеносных сосудов и т.д., при апоптозе клетка аккуратно саморазбирается на части, и соседние клетки используют ее фрагменты в качестве строительного материала.
Самоликвидации подвергаются и митохондрии - изучив этот процесс, Скулачев назвал его митоптозом. Митоптоз происходит, если в митохондриях образуется слишком много свободных радикалов. Когда количество погибших митохондрий слишком велико, продукты их распада отравляют клетку и приводят к ее апоптозу. Старение, с точки зрения Скулачева, - результат того, что в организме гибнет больше клеток, чем рождается, а отмирающие функциональные клетки заменяются соединительной тканью. Суть его работы - поиск методов противодействия разрушению клеточных структур свободными радикалами. По мнению ученого, старость - это болезнь, которую можно и нужно лечить, программу старения организма можно вывести из строя и тем самым выключить механизм, сокращающий нашу жизнь.
По мнению Скулачева, главная из активных форм кислорода, приводящих к гибели митохондрий и клеток - перекись водорода. В настоящее время под его руководством проходит испытания препарат SKQ, предназначенный для предотвращения признаков старения.
АДАПТАЦИОННО-РЕГУЛЯТОРНАЯ ТЕОРИЯ

Модель старения, разработанная выдающимся украинским физиологом и геронтологом В.В. Фролькисом в 1960-70-х гг., основана на широко распространенном представлении о том, что старость и смерть генетически запрограммированы. <Изюминка> теории Фролькиса состоит в том, что возрастное развитие и продолжительность жизни определяются балансом двух процессов: наряду с разрушительным процессом старения развертывается процесс <антистарения>, для которого Фролькис предложил термин <витаукт> (лат. vita - жизнь, auctum - увеличивать). Этот процесс направлен на поддержание жизнеспособности организма, его адаптацию, увеличение продолжительности жизни. Представления об антистарении (витаукте) получили широкое распространение. Так, в 1995 г. в США состоялся первый международный конгресс по этой проблеме.
Существенным компонентом теории Фролькиса является разработанная им генорегуляторная гипотеза, по которой первичными механизмами старения являются нарушения в работе регуляторных генов, управляющих активностью структурных генов и, в результате, интенсивностью синтеза закодированных в них белков. Возрастные нарушения генной регуляции могут привести не только к изменению соотношения синтезируемых белков, но и к экспрессии ранее не работавших генов, появлению ранее не синтезировавшихся белков и, как результат, к старению и гибели клеток.
В.В.Фролькис полагал, что генорегуляторные механизмы старения являются основой развития распространенных видов возрастной патологии - атеросклероза, рака, диабета, болезней Паркинсона и Альцгеймера. В зависимости от активации или подавления функций тех или иных генов и будет развиваться тот или иной синдром старения, та или иная патология. На основе этих представлений была выдвинута идея генорегуляторной терапии, призванной предупреждать сдвиги, лежащие в основе развития возрастной патологии.
РЕДУСОМНАЯ ТЕОРИЯ ОЛОВНИКОВА
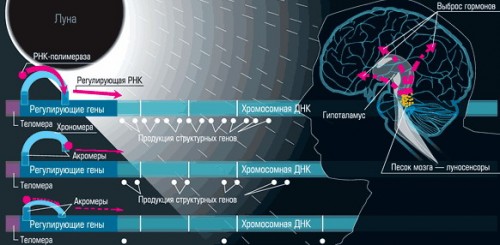
Покрытая белками линейная молекула ДНК редусомы - это копия сегмента хромосомной ДНК. гнезде. Подобно теломерной ДНК линейная ДНК редусомы с течением времени укорачивается. Поэтому крошечные редусомы прогрессирующе уменьшаются в размерах; отсюда и их название. Вместе с убылью ДНК в редусоме уменьшается и количество содержащихся в ней разных генов. Укорочение молекул редусомной ДНК (и вызванное этим изменение набора генов в редусомах, меняет с возрастом уровень экспрессии различных хромосомных генов и благодаря этому служит ключевым средством измерения биологического времени в индивидуальном развитии.
ТЕОРИЯ СОМАТИЧЕСКИХ МУТАЦИЙ

Среди современных теорий старения, основанных на предположении, что ДНК является основной мишенью повреждающих агентов в клетке, доминирует теория соматических мутаций, согласно которой старение - результат взаимодействия различных эндогенных и экзогенных повреждающих агентов с генетическим материалом клетки и постепенного накопления случайных мутаций в геноме соматических клеток. Повреждения ядерной и митохондриальной ДНК соматических клеток, такие, как точечные мутации, делеции и транслокации, приводят к активации или инактивации специфических генов, вовлеченных в регуляцию клеточного цикла и контроль роста. Накопление с возрастом таких мутаций в различных органах и тканях - основной фактор, определяющий развитие возрастной патологии, включая рак.
| <== предыдущая лекция | | | следующая лекция ==> |
| ТЕХНОЛОГИИ ИСКУССТВЕННОГО ИНТЕЛЛЕКТА | | | ВЫСОКОТЕМПЕРАТУРНОЕ ОБЕЗВРЕЖИВАНИЕ ГАЗОВ |
Дата добавления: 2017-01-08; просмотров: 2132;
Поиск по сайту
Узнать еще
- Arthropoda.Систематика.Блохи.Виды блох.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское и эпидемиологическое значение.Меры борьбы.
- Arthropoda.Систематика.Мошки,мокрецы,слепни,оводы.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское значение,меры борьбы.
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- Cоциально-политическое развитие Киевской Руси. Деятельность первых древнерусских князей.
- I.1.6 ОСНОВНОЕ УРАВНЕНИЕ КИНЕТИЧЕСКОЙ ТЕОРИИ ГАЗОВ. ЗАКОН РАВНОМЕРНОГО РАСПРЕДЕЛЕНИЯ ЭНЕРГИИ ПО СТЕПЕНЯМ СВОБОДЫ
- I.7.2 ПОСТУЛАТЫ РЕЛЯТИВИСТСКОЙ ТЕОРИИ
- II. Политическое развитие страны: реформы и их последствия
- II. Установление сегуната Токугава. Социально-экономическое развитие страны: циклы подъема и упадка

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
