МОЛЕКУЛЯРНВЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ
Генетическая информация, которую передает одно поколение клеток или организмов другому, включена преимущественно в ядре клетки. Ядро является обязательным структурным компонентом каждой эукариотической клетки.
Ядерная оболочка (кариолемма) интерфазного ядра состоит из двух элементарных мембран (наружной и внутренней), пространство между которыми называется перинуклеарным. Мембраны ядра имеют поровый комплекс. Через них идет обмен веществ между ядром и цитоплазмой, регуляция которого и является основной функцией ядерной оболочки. Наружная ядерная мембрана может переходить в стенки каналов зндоплазматической сети. На наружной ядерной мембране располагаются рибосомы. Внутренняя ядерная мембрана выстлана специальными белками (ламин и др.), которые образуют ядерную пластинку, обеспечивающую закрепление структур ядра (рис. 2.10).
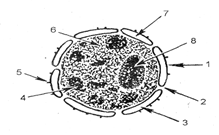
Рис. 1 Схема строения интерфазного ядра:
1 - наружная мембрана; 2 - пора; 3 - пери-нуклеарное пространство; 4 - глыбка хроматина; 5 - рибосома; 6 - кариолимфа; 7- внутренняя мембрана; 8 -ядрышко
Ядерный сок (кариолимфа) - однородный коллоидный раствор, заполняющий пространство между структурами ядра (хроматином и ядрышками). Он содержит воду, белки, нуклеотиды, АТФ, различные виды РНК, минеральные соли. Все ядерные белки синтезируются в цитоплазме и затем поступают в ядро. Кариолимфа осуществляет взаимосвязь ядерных структур и цитоплазмы клетки.
Хроматин - представляет собой дезоксирибонуклеопротеин (ДНП) - комплекс ДНК и гистоновых белков. Под световым микроскопом он выявляется в виде тонких нитей, глыбок, гранул. В процессе митоза, спирализуясь хроматин образует хорошо видимые, интенсивно окрашивающиеся структуры - хромосомы. Морфологически различают гетерохроматин и эухроматин. Эухромати менее конденсирован, чем гетерохроматин, и соответствует участкам хромосом, с которых идет считывание информации (транскрипция).
Белки ядра подразделяют на гистоновые и негистоновые. Гистоны - небольшие сильно основные молекулы белков. Они принимают непосредственное участие в упаковке молекулы ДНК, нейтрализуя отрицательно заряженные фосфатные группы ДНК своими положительно заряженными аминокислотными остатками. К группе негистоновых белков относятся структурные ядерные белки, многие ферменты факторы транскрипции, связанные с определенными участками ДНК и осуществляющие регуляцию генной активности.
Ядрышки - образования шаровидной формы (одно или несколько), состоящие из белков и РНК липидов, ферментов. Они не имеют мембраны. Ядрышки фрагментируются в начале деления клетки и восстанавливаются после его окончания. Образование ядрышек связано со вторичными перетяжками спутничных хромосом (ядрышковые организаторы). В области вторичных перетяжек локализованы гены, кодирующие синтез рибосомальной РНК (р-РНК), а в самих ядрышках происходит формирование субъединиц рибосом, которые затем выходят в цитоплазму через поры в ядерной оболочке.
Основные функции клеточного ядра:
▪ хранение и передача генетической информации;
▪ регуляция всех процессов жизнедеятельности клетки.
Дезоксирибонуклеиновая кислота (ДНК) — биополимер с молекулярной массой 10-100 млн, мономерами которого являются 4 типа нуклеотидов. В состав каждого нуклеотида входят пятиуглеродный сахар (пентоза) — дезоксирибоза, остаток фосфорной кислоты и одно из четырех азотистых оснований — аденин (А), гуанин (Г), цитозин (Ц) или тимин (Т). Два азотистых основания относятся к классу пуринов (А и Г) и два — к классу пиримидинов (Ц и Т). Нуклеотиды соединяются ковалентными (фосфодиэфирными) связями между фосфатной группой одного нуклеотида и дезоксирибозой другого. Сахарофосфатный остов полинуклеотида строится путем образования фосфодиэфирных мостиков между 3΄ - и 5΄ - положениями углерода молекул пентоз. К молекулам дезоксирибозы в качестве боковых радикалов присоединяются азотистые основания (рис. 3.5).
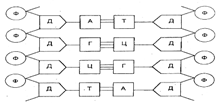
Рис. 3.5. Схема структуры участка молекулы ДНК:
Ф- остаток фосфорной кислоты; Д- дезоксирибоза; А, Г, Ц, Т- соответственно аденин, гуанин, цитозин, тимин (азотистые основания)
ДНК состоит из двух полинуклеотидных цепей, закрученных вправо вокруг одной оси с образованием двойной спирали. Цепи антипараллельны, т. е. направлены в противоположные стороны, так что 3'-конец одной цепи располагается напротив 5'-конца другой. Каждая цепь состоит из сахарофосфатного остова, вдоль которого перпендикулярно к длинной оси двойной спирали располагаются основания. Находящиеся друг против друга основания двух цепей двойной спирали соединяются водородными связями между пуриновыми и пиримидиновыми основаниями строго комплементарно: аденин — только с тимином (две связи), а гуанин — с цитозином три связи) (рис. 3.6).
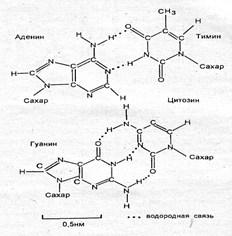
Рис. 3.6. Спаривание азотистых оснований:
аденина с Тимином, гуанина с цитозином
Расстояние между сахарофосфатными стовами двух цепей постоянно и равно расстоянию, занимаемому парой оснований, т. е. одним пурином и одним пиримидином. Вдоль оси молекулы соседние пары оснований рас-полагаются на расстоянии 0,34 нм одна от другой. Полный борот спирали — 3,4 нм, т. е. 10 пар оснований (рис. 3.7).
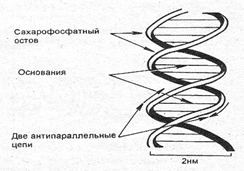
Рис. 3.7. Схема двухцепочечной структуры молекулы ДНК
ДНК является хранителем генетической информации во всех клетках про- и эукариот. У некоторых доклеточных форм (вирусы и бактериофаги) эту функцию выполняет молекула НК. Основная масса ДНК клетки сосредоточена в ядре 99%), небольшое ее количество находится в ДНК-содержащих органоидах (митохондрии, пластиды).
Рибонуклеиновая кислота (РНК) также является полинуклеотидом, но в отличие от ДНК ее молекула, как правило, состоит из одной цепочки. В состав нуклеотидов РНК входят пятиуглеродный сахар — рибоза и азотистые основания аденин, урацил (вместо тимина), гуанин и цитозин. Различают три вида РНК: информационную (и-РНК), транспортную (т-РНК) ирибосомал ьную (р-РНК).
Уровни упаковки
Дата добавления: 2016-11-26; просмотров: 2674;
Поиск по сайту
Узнать еще
- II. ОСНОВЫ МОЛЕКУЛЯРНОЙ ФИЗИКИ И ТЕРМОДИНАМИКИ
- V. ЛОГИЧЕСКИЕ ОСНОВЫ ВЫЧИСЛИТЕЛЬНОЙТЕХНИКИ
- XIV. ОСНОВЫ МАССОПЕРЕДАЧИ
- ІІ.5.2. Основы процесса фракталь-ного расширения квадрата
- Анатомо - физиологические основы массажа
- Арифметические основы ЭВМ
- Атомное ядро и основы ядерной энергетики
- Билет № 20 (2) Понятие «ткань». Подготовка основы и утка к ткачеству.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
