КРАТКАЯ ХАРАКТЕРИСТИКА МИКРООРГАНИЗМОВ И ОСНОВНЫЕ МЕТОДЫ МИКРОБИОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ
ГЛАВА 1
МОРФОЛОГИЯ МИКРООРГАНИЗМОВ.
ХАРАКТЕРИСТИКА ОТДЕЛЬНЫХ ГРУПП
Морфология микроорганизмов изучает форму и строение их клеток, способы передвижения и размножения. Микроорганизмы различаются по внешнему виду и по размерам. Строение клеток микроорганизмов также различно, в связи с чем они относятся к различным систематическим группам.
Все живые организмы на Земле , имеющие клеточное строение, делят на два надцарства: прокариоты и эукариоты. Это деление живых организмов основано главным образом на особенностях строения ядерного аппарата. В клетках прокариот ядро отсутствует. Ядерный аппарат их представлен молекулой ДНК, расположенной в ядерной зоне непосредственно в цитоплазме. Клетки эукариот имеют ядро, отделенное от цитоплазмы двойной ядерной мембраной.
БАКТЕРИИ
Известно около 4000 видов бактерий. Их разнообразие особенно выражено в отношении физиолого-биохимических свойств. В определенной степени оно проявляется и в морфологии.
Величина клеток различных бактерий сильно варьирует. Размеры многих бактериальных форм находятся в пределах 0,5-10 мкм. Однако величина ряда бактерий не укладывается в эти границы. Среди них есть немало относительно крупных форм, есть и крайне мелкие формы. Значительной длины достигают, например, нитчатые бактерии рода Beggiatoa - до 60 мкм и более и Saprospira - до 500 мкм. Это одни из наиболее крупных бактерий. Гигантские формы встречаются среди спирохет: длина некоторых достигает 500 мкм. Мельчайшие из известных организмов клеточного строения – микоплазмы. Размеры отдельных форм микоплазм не превышают 0,1-0,2 мкм, что лежит на границе или даже за пределами разрешающей способности светового микроскопа. У одного и того же вида бактерий размеры клеток могут в большей или меньшей степени варьировать в зависимости от возраста культур и (или) от условий культивирования. У многих бактерий особенно заметно меняется длина клетки. Диаметр клеток является более устойчивым признаком.
Основная масса бактерий - одноклеточные организмы. Но нередко клетки после деления не расходятся и образуют сочетания различной формы, которая определяется расположением делящей перегородки. Эти сочетания не равноценны многоклеточным организмам, так как каждая клетка в них автономна и может существовать самостоятельно после отделения от остальных клеток.
Бактерии, за исключением микоплазм, имеют определенную форму клетки. У большинства бактерий она поддерживается благодаря прочной (ригидной)
клеточной стенке. Клеточная стенка спирохет эластична, и их извитая форма поддерживается с помощью аксиальных фибрилл, расположенных под клеточной стенкой. Форма клетки многих бактерий отличается постоянством и сохраняется в течение всей жизни. Но есть бактерии, у которых наблюдается более или менее выраженный плеоморфизм. Нередко он отражает стадии цикла развития микроорганизма. В этом случае обнаруживается упорядоченное, регулярное чередование определенных форм. Изменения морфологии могут происходить и под влиянием условий культивирования. Полиморфность микоплазм связана с отсутствием у них клеточной стенки.
Морфологические типы бактерий по сравнению с высшими организмами немногочисленны. Клетки значительной части бактерий имеют сферическую, цилиндрическую или спиралевидную форму. Существует обширная группа ветвящихея бактерий, сравнительно небольшое количество нитчатых форм и бактерий, образующих выросты (простеки).
Сферические бактерии - кокки. Под микроскопом они имеют форму шара. Многим коккам свойственно образование различных сочетаний (рис. 2). Кокки, делящиеся в одной плоскости и одном направлении, могут образовывать пары (диплококки) или цепочки (стрептококки) клеток. Когда деление происходит равномерно в двух взаимно перпендикулярных плоскостях, возникают группы

Рисунок 2. Сочетания кокков: 1 – диплококки; 2 – стрептококки; 3 – тетракокки и сарцины; 4 – стафилококки и микрококки
из четырех клеток – тетракокки, а если в трёх, то образуют пакеты правильной формы – сарцины. При неравномерном делении в нескольких плоскостях наблюдаются скопления неправильной формы, напоминающие гроздь винограда. Они свойственны представителям стафилококков и микрококков. Микрококками часто называют и одиночные шаровидные клетки.
Под влиянием различных факторов среды некоторые кокки могут превращаться в овальные, конические и эллипсоидные клетки.
Цилиндрические (палочковидные) бактерии под микроскопом имеют вид палочек. Это одна из наиболее многочисленных групп бактерий. Разные виды могут заметно отличаться друг от друга размерами клеток. Одной из самых крупных палочковидных бактерий является Васilllus megaterium. Ее длина 5-10 мкм, поперечник около 1 мкм. К наиболее коротким относятся риккетсии, размеры которых могут быть всего 0,3 Х 1,0 мкм. В тех случаях, когда длина лишь ненамного превышает диаметр клетки, палочки трудно отличить от кокков. Концы палочек бывают прямыми, округлыми или заострёнными (рис. 3).

Рисунок 3. Палочковидные бактерии: 1 - Pseudomonas aeruginosa; 2 - Bacillus mycoides; 3 – Васillus megaterium; 4 – Cytophaga
Палочковидные бактерии нередко образуют пары или цепочки клеток. Парные сочетания клеток наблюдаются, например, у определенных видов рода Pseudomonas, длинные цепочки можно увидеть в культуре Bacillus mucoides. Для ряда палочковидных бактерий характерен выраженный плеоморфизм.
Изменение формы, связанное с развитием бактерий, наблюдается у видов Azotobacter и Rhizobium; у миксобактерий и риккетсий. Так уже в молодой культуре азотобактера можно видеть клетки не только палочковидной, но и овальной или кокковидной формы. Они часто соединяются попарно или образуют скопления, а иногда цепочки из 4 и более клеток. В старых культурах преобладают крупные округлые, неправильной формы покоящиеся клетки-цисты .Риккетсии, помимо коротких палочек длиной 1-1,5 мкм могут быть представлены кокками диаметром менее 0,5 мкм, длинными палочками - 3-4 мкм, или причудливо изогнутыми нитями, длина которых достигает 40 и более микрометров. Есть бактерии, у которых изменение формы клетки связано со спорообразованием.
В неблагоприятных условиях в культурах многих палочковидных бактерий возникают различные дегенеративные формы с признаками лизиса, гранулированием содержимого, большими вакуолями и др. Это можно наблюдать, например, в культуре Bacillus megaterium (рис. 3).
Извитые одноклеточные бактерии бывают трех типов: вибрионы, спириллы и спирохеты. Вибрионы выглядят как слегка изогнутые палочки, похожие на запятую (рис. 4). В пpoцecce развития некоторые из них могут менять форму клеток. Так, паразиты бактерий Bdellovibrio при развитии в периплазматическом пространстве клетки хозяина становятся спиралевидными. Впоследствии спиралевидные клетки распадаются на вибрионы. Спириллы похожи либо на латинскую букву S, либо на штопор, т. е. имеют несколько правильных завитков. Спирохеты имеют вид тонких спиралевидных клеток с многочисленными завитками и петлями. Длина клеток спирохет превышает толщину в 5 - 200 раз. Спирохеты нередко образуют аномальные формы - гранулы, сфероиды и др.

Рисунок 4 Извитые формы: 1 – вибрионы; 2 – спириллы; 3 – спирохеты
Бактерии, образующие выросты (простеки). Основную часть этой группы составляют бактерии, у которых простеки - это выпячивания клеточного содержимого, окруженного клеточной стенкой цитоплазматической мембраной и не отделенного от клетки перегородкой. У одних бактерий, например у видов рода Hyphomicrobium, образование выростов связано с размножением. Клетки представителей этого рода чаще имеют вид палочек с заостренными концами, но бывают также овальной, яйцеобразной или бобовидной формы. Нитевидные выросты образуются на одном или обоих полюсах клетки. Bыросты могут ветвиться, давая гифоподобные структуры. На конце каждой ветви формируется почка, являющаяся дочернеи клеткой. Иногда созревшие почки не отделяются от материнской клетки и тоже образуют выросты и почки. Тогда возникает скопление гиф и клеток (рис. 5).
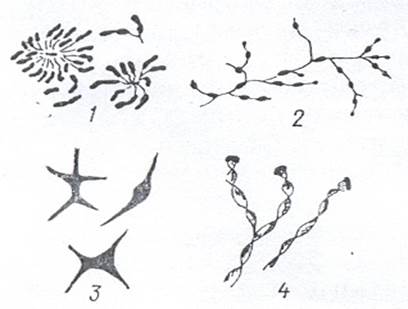
Рисунок 5 Бактерии, образующие выросты: 1 – Caulobacter; 2 – Hyphomicrobium; 3 – Ancalomicrobium; 4 – Gallionella
У других бактерий простеки не имеют отношения к размножению. К таким бактериям принадлежат, например, виды рода Caulobacter и Ancalomicrobium. Клетки Caulobacter – это слегка изогнутые палочки с одним полярным жгутиком. Сравнительно короткий вырост - стебелек возникает на одном полюсе клетки. На конце стебелька имеется небольшое утолщение из липкого материала - фиксатор. С его помощью клетки прикрепляются к какому-либо субстрату, а иногда друг к другу. В последнем случае образуются характерные скопления. У видов Аnсаlomicrobium на клетке неправильной формы возникает несколько простеков - от 2 до 8. Клетка приобретает причудливый звездообразный вид.
Иногда к стебельковым относят бактерии, образующие слизистые придатки, не связанные с цитоплазмой клетки. Это, например, виды Gallionella, бобовидные клетки которой выделяют с вогнутой стороны слизь в виде тонкой нити. Под микроскопом такая нить выглядит как спирально изогнутая лента.

Рисунок 6. Нитчатые бактерии: 1 - Beggiatoa; 2 - Thiothrix; 3 - Saprospira; 4 - Simonsiella; 5 - Caryophanon; 6 - цианобактерии класса Hormogoneae; 7 - Leptothrix; 8 - Sphaerotilus; 9 - Crenothrix
Нитчатые бактерии. Это сравнительно небольшая группа многоклеточных организмов. Они представляют собой цепочки (трихомы) из цилиндрических, овальных или дисковидных клеток. Типичными представителями нитчатых форм являются бактерии родов Beggiatoa и Thiothrix (рис.6). Их нити имеют равную толщину на всем протяжении. Трихомы видов Thiothrix собраны в пучки и прикрепляются основанием к субстрату. Нити Leucothrix, подобно Thiothrix, большей частью также растут пучком, прикрепляясь к твердой поверхности, но, в отличие от Thiothrix, они сужаются к концу.
Трихомы видов Saprospira скручены в виде спирали, а у видов Simоnsiella они уплощены и похожи на ленты. У видов Caryophanoп поперечные клеточные стенки большинства составляющих нити клеток не сплошные, так как их формирование отстает от роста трихома. Нитчатые бактерии относятся к крупным микроорганизмам. Так, длина нитей некоторых представителей рода Caryophanon достигает 40 мкм, а толщина 4 мкм. Нити зеленых бактерий группы Chloroflexus могут иметь длину 300 мкм. Особенно длинные трихомы образуют, как уже отмечалось, виды Beggiatoa и Saprospira (до 500 мкм).
Ветвящиеся бактерии. К этой многочисленной группе относятся истинные актиномицеты, нокардии, микобактерии, коринеподобные бактерии и ряд других организмов. Истинные актиномицеты имеют сильноразветвленный мицелий, сохраняющийся в течение всей жизни, что делает их внешне сходными с мицелиальными грибами (рис. 7). Однако общая длина нитей актиномицетов обычно не превышает нескольких миллиметров, а толщина составляет всего 0,5-1,5 мкм, тогда как длина грибного мицелия достигает нескольких сантиметров, а диаметр может быть около 50 мкм. У представителей рода Streptomyces в мицелии образуются перегородки, но их мало, поэтому составляющие его клетки в основном многоядерные. Мицелий большинства актиномицетов лишен перегородок, и этим он напоминает многоядерный несептированный мицелий фикомицетов.
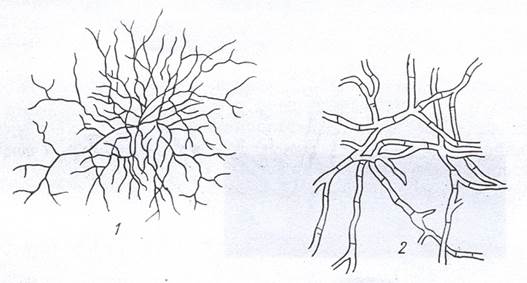
Рисунок 7. Мицелий актиномицета (1) и гриба (2) при одинаковом увеличении
У нокардий и микобактерий мицелиальный тип развития имеет временный и часто ограниченный характер. Виды рода Nocardia образуют обильный, недифференцированный мицелий на начальных стадиях развития. В дальнейшем он распадается на палочковидные или сферические фрагменты.
Микоплазмы. Это довольно большая группа бактерий, у которых нет клеточной стенки. Поэтому они очень полиморфны. В культуре одного вида можно одновременно обнаружить мелкие зерновидные образования, кокковидные, эллипсовидные, грушеобразные, дисковидные, палочковидные и даже разветвленные и неразветвленные нитевидные формы (рис. 8). Размеры крупных клеток микоплазм достигают 10 мкм, а величина мелких структур не превышает 0,1 мкм.
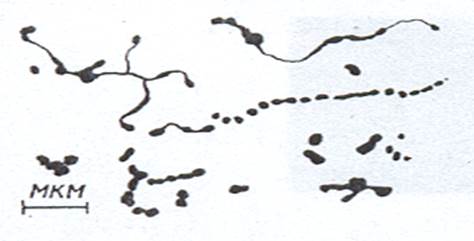
Рисунок 8. Микоплазмы. Схема электронной микрофотографии
Большинство бактерий размножаются путем бинарного поперечного изоморфного деления. Такой способ размножения свойствен коккам, многим палочковидным формам и вибрионам, спириллам, спирохетам, некоторым нитчатым бактериям. Клетки основной массы бактерий делятся в одной плоскости. У многих кокков деление происходит в нескольких плоскостях. Расходящиеся после деления клетки большинства бактерий располагаются одна за другой или беспорядочно, а у видов Arthrobacter и Corynebacteriuт под углом друг к другу. Если после деления клетки не расходятся, то наблюдается образование различных скоплений клеток - пар, цепочек, пакетов и другие. В ряде случаев имеет место неравномерное деление. Фрагментацией мицелия или его рудиментов на палочки и кокки размножаются, например, виды Nocardia и Mycobacteriuт. Размножение распадом нитей на участки наблюдается у Beggiatoa и Saprospira. Две неодинаковые клетки - одна подвижная со жгутом, но без простеки, а другая неподвижная без жгутика, но со стебельком - образуются при делении клеток Caulobacter (рис. 9). К делению способны только неподвижные клетки с простекой.
Некоторые бактерии (виды Hyphoтicrobiuт и Rhodopseudoтonas, Ancaloтicrobiuт и др.) размножаются почкованием. У Rhodopseudoтoпas и Aпcaloтicгobiuт почки формируются прямо на поверхности клеток, а у Hyphoтicrobiuт - на концах гиф .
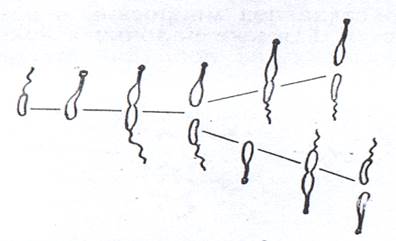
Рисунок 9. Схема роста и деления клеток Caulobacter
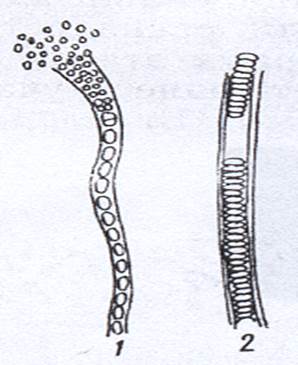
Рисунок 10. Гонидии (1) и гормогонии (2) нитчатых бактерий
У бактерий известны и более сложные способы размножения. Нитчатые цианобактерии класса Chaтaesiphoneae и бактерии родов Thiothrix, Caryophanon, Sphaerotilus, Leptothrix, Leucothrix размножаются с помощью специальных репродуктивных одиночных подвижных клеток - гонидий (рис. 10), которые образуются в результате многократного деления концевых клеток нити. Подвижность гонидий связана с наличием у них жгутиков. Для нитчатых цианобактерий класса Horтogoпeae характерно размножение гормогониями. Это короткие цепочки, возникающие, как и гонидии, при делении клеток нити. Они не имеют жгутиков и перемещаются скольжением благодаря выделению слизи. Размножение гормогониями наблюдается также у видов Leucothrix.
Актиномицеты размножаются главным образом подвижными или неподвижными спорами (конидиями). Конидии располагаются поодиночке или цепочками, непосредственно на мицелии, на концах спороносящих гиф - спорангиеносцах (спорангиофорах) или в специальных органах спороношения - спорангиях. Спорангиеносцы (и соответственно цепочки спор) разных видов различаются между собой. Они могут быть длинными или короткими, прямыми, волнистыми или спиралевидными; иметь последовательное, супротивное или мутовчатое расположение (рис. 11). Спорангии бывают сферической или неправильной формы (рис. 12), в них формируются эндогенные споры.
Существует немало бактерий, которые могут размножаться несколькими способами. Например, представители рода Rhizobiuт размножаются делением и почкованием, актиномицеты - спорами и кусочками вегетативного мицелия. Нитчатые цианобактерии размножаются гонидиями или гормогониями, а также путем распада трихома на отдельные участки, бактерии рода Chloroflexus - бинарным делением и участками нити. Caryophanon и Sphaerotilus - с помощью гонидий и поперечным изоморфным делением трихома, Leucothrix гонидиями и гормогониями. У микоплазм можно наблюдать бинарное деление, фрагментацию нитей и крупных клеток до кокков, а также процесс, напоминающий почкование.

Рисунок 11 Форма воздушных спороносцев у актиномицетов
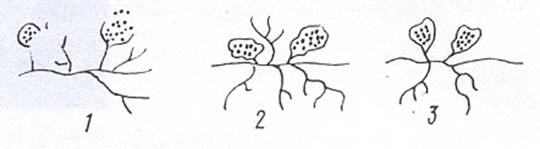
Рисунок 12. Спорангии актиномицетов: 1 – Actinoplanes; 2 – Amorphosporangium; 3 - Spirillospora
Многие бактерии неподвижны. Неподвижными являются почти все кокки, более 50% палочковидных бактерий, почкующиеся и ветвящиеся бактерии, значительная часть нитчатых форм, риккетсии, микоплазмы. Способностью к движению обладает примерно 1/5 часть бактерий. Подвижность большинства из них обусловлена наличием специальных локомоторных структур - жгутиков. Жгутики обнаруживаютсяу некоторых кокков (отдельные представители рода Methylococcus), ряда палочковидных бактерий (виды Bacillus, Clostridiuт, Pseudoтoпas, Rhizobium, Azotobacter, Escherichia и др.), у вибрионов и спирилл, у нитчатых бактерий рода Caryophanon. У бактерий некоторых групп специальные репродуктивные клетки со жгутиками появляются только в определенной стадии развития. Это подвижные клетки каулобактерий, гонидии большинства нитчатых организмов, споры (конидии) некоторых актиномицетов (виды Actinoplaпes и Geoderтatopftilus).

Рисунок 13. Типы жгутикования у бактерий: 1 – монотрихиальное; 2 – лофотрихиальное; 3 – латеральное; 4 – амфитрихиальное; 5 – перитрихиальное; 6 – «смешанное» полярно - перитрихиальное
Жгутики берут начало под цитоплазматической мембраной и через поры мембраны и клеточной стенки выходят наружу. У разных бактерий длина жгутиков колеблется от 3 до 20 мкм, толщина - от 10 до 20 им, а их число - от 1 до 100. Жгутики могут быть расположены монополярно, биполярно, вдоль боковой или по всей поверхности клетки (рис. 13). Клетки некоторых бактерий имеют одновременно два разных набора жгутиков: полярные и перитрихиальные, различающиеся по длине и толщине.
Наличие, число, размеры и расположение жгутиков имеют диагностическое значение. Например, виды рода Vibrio снабжены одним полярным жгутиком, у Selenoтonas один жгутик прикрепляется сбоку. Для представителей рода Pseudoтonas характерно монотрихиалыюе или лофотрихиальное монополярное жгутикование, а для спирилл лофотрихиальное моно- и биполярное. Перитрихиальное расположение жгутиков свойственно видам Clostridium, Escherichia, Rhizobium, Саryophanon и др. Нередко в пределах одного рода бактерий обнаруживаются подвижные и неподвижные виды, а у подвижных форм может быть разный тип жгутикования. Так, у подвижных представителей рода Bacillus жгутики расположены латерально или перитрихиально.
Активное движение большинства бактерий, обладающих жгутиками, возможно только в жидкой среде. Однако некоторые бактерии - перитрихи могут передвигаться и по твердому субстрату. К ним относится, например, Proteus vulgaris, который довольно быстро распространяется по поверхности. влажной агаризованной среды, образуя обширный тонкий налет. Движение жгутиконосных бактерий наблюдается преимущественно в молодых культурах. С возрастом клетки постепенно теряют жгутики и становятся неподвижными, хотя и сохраняют жизнеспособность.
К подвижным формам относятся спирохеты, миксобактерии, многие нитчатые цианобактерии и флексибактерии, не имеющие жгутиков.
Они способны передвигаться по твердому или полутвердому субстрату
путем скольжения. Спирохеты могут перемещаться и в жидкой среде
вращательными, легкими волнообразными движениями. Скользящее
движение обусловлено, возможно, неравномерным выделением слизи
через поры клеточной стенки. Подвижность спирохет и некоторых миксобактерий (виды Myxococcus) связывают также с сокращением аксиальных микрофибрилл, расположенных под клеточной стенкой (у спирохет) или под цитоплазматической мембраной (у миксобактерий).
К покоящимся формам бактерий относятся эндоспоры, цисты, акинеты. Они позволяют клетке более или менее длительное время переносить неблагоприятные условия. В условиях, подходящих для роста, покоящиеся формы развиваются в обычную вегетативную клетку.
Эндоспоры. Способностью образовывать эндоспоры обладают палочковидные бактерии, относящиеся к родам Bacillus, Clostridiuт и
Desulfotoтaculuт, а также некоторые кокки (род Sporosarcina) и термофильные актиномицеты рода Therтoactinoтyces. Спорообразование представляет собой сложный процесс дифференцировки, начинающийся в культуре, когда она переходит в стационарную фазу роста и когда создаются условия, индуцирующие его. Эти условия весьма разнообразны: дефицит питательных веществ в среде, накопление продуктов метаболизма, изменение кислотности среды, температуры и др. В результате внутри вегетативной клетки образуется новая клетка - эндоспора, полностью отличающаяся от материнской по структуре, химическому составу и физиологическим свойствам. Эндоспоры одеты толстыми многослойными труднопроницаемыми покровами и имеют очень низкое содержание воды, поэтому при микроскопическом исследовании их легко узнать по высокой светопреломляющей способности.
Форма клеток многих бактерий в процессе спорообразования не меняется. Эндоспора локализуется в центре клетки, эксцентрально или (и) терминально, что зависит от вида бактерий. Это так называемый бациллярный тип спорообразования (рис. 14, 1). У ряда бактерий середина клетки при формировании споры несколько расширяется, и клетка приобретает вид челнока или веретена. Спора располагается в утолщенной части - в центре клетки или эксцентрально (рис. 14, 2). Это - клостридиальный тип спорообразования. У некоторых бактерий клетка при спорообразовании сильно расширяется и округляется на одном конце, становясь похожей на барабанную палочку. Спора локализуется в расширенном конце (рис. 14, 3). Такой тип спорообразования называется плектридиальным. Бациллярный тип спорообразования свойствен многим представителям рода Bacillus, клостридиальный и плектридиальный - в основном видам рода Clostridiuт. Нередко в культуре одного вида этого рода встречаются одновременно и клостридиальные и плектридиальные формы.

Рисунок 14. Типы образования эндоспор у бактерий: 1 – бациллярный; 2 – клостридиальный; 3 – плектридиальный
Эндоспоры бывают округлой, овальной или эллипсовидной формы. Их оболочка может быть гладкой или с выростами. Диаметр эндоспор ряда бактерий значительно превышает поперечник клетки. Тип спорообразования, а также форма, размеры и расположение эндоспоры в вегетативной клетке используются для диагностики бактерий.
В каждой вегетативной клетке формируется, как правило, только одна эндоспора. После созревания эндоспоры освобождаются вследствие лизиса материнских клеток и переходят в стадию покоя. Эндоспоры чрезвычайно устойчивы к различным неблагоприятным факторам и могут сохранять жизнеспособность в течение многих лет, пока не попадут в условия, способствующие их прорастанию.
Спорообразование - не обязательная стадия развития бактерий. Можно создать такие условия, в которых клетки не будут переходить к формированию спор.
Цисты обнаруживаются у миксобактерий, риккетсий, представителей родов Azotobacter, Bdellovibrio, Arthrobacter. Их образование происходит обычно на поздних стадиях развития бактерий и связано с неблагоприятными условиями культивирования - исчерпанием питательного субстрата, загрязнением среды вредными продуктами обмена, высушиванием и т. д. Цисты можно увидеть только в старых культурах.
Цисты бывают сферическими, овальными, неправильно округлыми или в виде сильно укороченных палочек. Чаще всего они крупнее вегетативных клеток . Иногда же по форме и размерам цисты почти не отличаются от них. У большинства бактерий цисты имеют утолщенную клеточную стенку и уплотненную цитоплазму, поэтому они сильнее преломляют свет, чем вегетативные клетки. Цисты устойчивее вегетативных клеток к неблагоприятным факторам, но уступают в этом эндоспорам.
Акинеты свойственны определенным видам нитчатых цианобактерий. Это крупные толстостенные клетки (рис. 15), возникающие либо из одной вегетативной клетки, либо путем слияния многих клеток. У некоторых цианобактерий акинеты обнаруживаются всегда и являются, вероятно, обязательной стадией развития, у других они образуются только в неблагоприятных условиях.
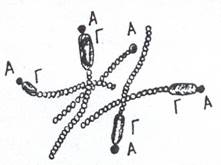
Рисунок 15. Акинеты (а) и гетероцисты ( Г) нитчатой цианобактерии Cylindrospermum
Клетки всех бактерий, за исключением микоплазм, покрыты снаружи клеточной стенкой, толщина которой у разных видов колеблется в пределах 0,01-0,04 мкм. В соответствии с различиями в химическом составе клеточных стенок и их ультраструктуре, выражающимися в неодинаковой способности клеточных стенок удерживать красители трифенилметанового ряда с йодом, прокариотные микроорганизмы делятся на две группы. К одной относятся бактерии, в клетках которых комплекс, образуемый кристаллическим или генциановым фиолетовым и йодом, не обесцвечивается при последующей обработке спиртом. К другой группе принадлежат бактерии, не обладающие свойством удерживать краситель и обесцвечивающиеся при обработке спиртом. Этот способ дифференциальной окраски бактерий был предложен в 1884 году датским физиком Христианом Грамом. Бактерии, которые способны окрашиваться по Граму, называются грамположительными, а не способные окрашиватьсся - грамотрицательными. К первой группе относится большинство кокковых форм, спорообразующие палочковидные бактерии родов Bacillus и Clostridium, нитчатые бактерии Сагуорhanon, ветвящиеся бактерии. Ко второй приyадлежат различные палочковидные бактерии, не образующие эндоспор (роды Pseudoтonas, Escherichia и др.), простекобактерии, миксобактерии, риккетсии, многие нитчатые формы, спириллы, спирохеты, некоторые кокки и др. Химический состав и строение клеточных стенок грамотрицательных микроорганизмов значительно сложнее, чем грамположительных.
С особенностями химического состава клеточных стенок связывают и кислотоустойчивость микобактерий. Она выражается в способности клеток, фиксированных и окрашенных при подогревании карболовым фуксином, прочно удерживать окраску после обработки раствором минеральной кислоты или подкисленным спиртом .
Определенными способами, например, под действием лизоцима, бактериальные клетки могут быть лишены клеточных стенок. В таком виде они способны существовать только в изотонической питательной среде.
Клеточная стенка многих бактерий снаружи может быть окружена слизистым слоем - капсулой. Капсулы бывают полисахаридной, иногда гликопротеидной или полипептидной природы. Капсулы толщиной менее 0,2 мкм, неразличимые в световом микроскопе, называют микрокапсулами. Капсула и клеточная стенка являются поверхностными структурами бактериальной клетки, к которым относят также жгутики и обнаруживаемые у многих подвижных и неподвижных бактерий ворсинки (фимбрии, пили). Ворсинки короче и тоньше большинства жгутиков – их длина 3-4 мкм, диаметр 4-35 нм. Число ворсинок у разных бактерий бывает от нескольких единиц до многих тысяч. К подвижности бактерий они, по-видимому, не имеют отношения. Капсулы и ворсинки не являются необходимыми клеточными структурами. Бактерии нормально функционируют и без них.
Обязательной структурой любой клетки является цитоплазматическая мембрана, которая отделяет цитоплазму от клеточной стенки. Толщина мембраны. 5-10 нм. При нарушении ее целостности клетки утрачивают жизнеспособность. Цитоплазма ряда бактерий пронизана мембранными структурами, которые являются производными цитоплазматической мембраны. У гетеротрофных бактерий их называют мезосомами. Они имеют вид пластинок (ламелл), пузырьков (везикул) или трубочек. Мезосомы могут быть расположены в зоне клеточного деления, вблизи нуклеотида и на периферии клетки, недалеко от цитоплазматической мембраны. У грамположительных бактерий мезосомальные структуры развиты в большей степени, чем у грамотрицательных. У фототрофных бактерий мембранные образования в виде пузырьков называют хроматофорами, а уплощенной формы - тилакоидами. Есть бактерии, у которых мембранная система не обнаруживается.
Определенную область в цитоплазме бактериальной клетки занимает нуклеоид. Он состоит из одной двойной спирально закрученной нити ДНК, замкнутой в кольцо. Ядерный аппарат прокариот не имеет ядрышка и не отделен от цитоплазмы мембраной. Через мезосомы нуклеоид связан с цитоплазматической мембраной. В период интенсивного деления в клетках ряда бактерий (Escherichia соli, Oscillatoria атоеnа) можно обнаружить несколько нуклеоидов.
В цитоплазме бактерий в свободном виде или в связи с мембранными структурами находятся рибосомы. Они имеют константу седиментации 70S, их размеры колеблются в пределах от 15 до 30 нм. Число рибосом может быть от 5 до 50 тыс., что зависит от возраста клетки и условий культивирования. Рибосом больше в молодых клетках.
В клетках различных бактерий часто обнаруживаются включения
запасных веществ. Это полисахариды, липиды, полифосфаты, сера. Они накапливаются при избытке тех или иных питательных веществ в окружающей среде, а расходуются при голодании. Из резервных полисахаридов особенно распространены глюканы: гликоген, крахмал и крахмалоподобное вещество - гранулёза. Они выявляются в клетках спорообразующих бактерий родов Bacillus и Сlоstгidium, а также у пурпурных бактерий и др. Полисахариды откладываются в цитоплазме равномерно или в виде гранул. Запасные липиды 6актерий представлены полиэфиром 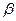 - оксимасляной кислоты и восками. Полиоксибутират накапливается на средах с избытком углерода у многих
- оксимасляной кислоты и восками. Полиоксибутират накапливается на средах с избытком углерода у многих
бактерий: видов Bacillus, Pseudoтanas, Spirilluт, Azotobacter, Sphaerotilus и др. Он обнаружен только у прокариот. Воска - эфиры высокомолекулярных жирных кислот и спиртов характерны для микобактерий. Полисахариды и липиды служат хорошим источником углерода и энергии для клетки.
В условиях, препятствующих синтезу нуклеиновых кислот, у многих бактерий создается резерв фосфора в виде гранул полифосфатов. Впервые они были описаны у Spirillит volutans, поэтому их назвали волютином. Эти образования называют также метахроматиновыми зернами, так как они проявляют метахроматический эффект: приобретают красную окраску при обработке синим красителем.
Отдельные виды спорообразующих бактерий (Bacillus thuringiensis, Bacillus cereus, Bacillus popilliae и др.) в определенных условиях образуют в клетках кристаллы белковой природы, которые имеют правильную бипирамидальную форму и расположены непосредственно около споры. Их называют параспоральными тельцами.
Некоторые бактериальные структуры и включения, сильно преломляющие свет (эндоспоры, аэросомы, отложения полиоксибутирата и серы), хорошо заметны в световом микроскопе без специальной обработки. Часть структур (жгутики, клеточная стенка, нуклеоид, волютин и др.) можно выявить с помощью светооптического микроскопа только после окрашивания соответствующими красителями. Ряд структурных элементов бактерий - микрокапсулы, ворсинки, мезосомы, рибосомы и др. различимы только 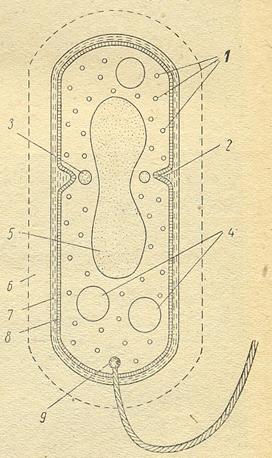
Рисунок 16. Схема строения бактериальной клетки: 1 – рибосомы, 2 – начавшееся образование поперечной перегородки, 3 и 4 – запасные отложения, 5 – ядерный район, 6 – капсула, 7 – стенки клетки, 8 – протоплазматическая мембрана, 9 – зерно, от которого начинается жгутик
в электронном микроскопе (рис.16).
ДРОЖЖИ
В группу дрожжей объединяются грибные организмы, которые в ростовой фазе существуют преимущественно в виде отдельных клеток. Примерно 2/3 дрожжей относится к классу сумчатых грибов(Ascoтycetes). Дрожжи есть также в классе базидиомицетов (Basidioтycetes) и дейтеромицетов - несовершенных грибов (Deuteroтycetes - Fungi iтperfecti).
Клетки разных видов дрожжей морфологически весьма разнообразны. Они бывают круглые, овальные, цилиндрические, яйцевидные, лимоновидные, колбовидные, треугольные, стреловидные и серповидные. Дрожжевые клетки значительно крупнее бактериальных. Длина их варьирует у разных видов от 2 до 20, иногда до 50 мкм, ширина от 1,5 до 10 мкм. К числу наиболее крупных дрожжей относятся, например, представители родов Saccharoтyces и Lipoтyces, наиболее мелкими являются виды Pichia и Наnsenula.
Как уже отмечалось, дрожжи существуют в основном в одноклеточной форме. Однако в культурах многих видов наблюдаются скопления разных размеров и очертаний, а некоторые дрожжи на определенной стадии развития могут образовывать мицелиальные структуры (рис. 17). Дрожжи являются неподвижными организмами.
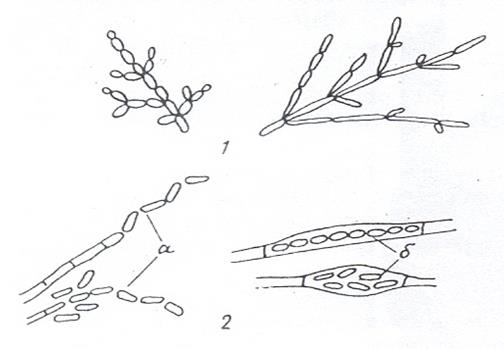
Рисунок 17. Мицелиальные формы дрожжей: 1 – псевдомицелий; 2 – истинный мицелий; а – артроспоры; б – эндоспоры
Дрожжи размножаются разными способами: вегетативно, бесполыми спорами и половым путем. Наиболее распространенным способом вегетативного размножения является почкование (рис. 18, 1). К почкующимся дрожжам относятся, например, виды родов Saccharoтyces и Lipoтyces. Если при почковании вновь возникающие клетки не отделяются друг от друга, то образуется псевдомицелий. Он отличается тем, что в зоне перегородок между клетками имеются перетяжки (рис. 18, 1). Образование псевдомицелия особенно характерно для дрожжей рода Candida.
Размножение делением (рис. 18, 2) встречается реже. Оно свойственно, например, видам Schizosaccharoтyces. Когда не расходятся делящиеся клетки, формируется истинный мицелий. Между клетками истинного мицелия четко различается перегородка, но нет перетяжек (рис. 18, 2). У ряда дрожжей (определенные виды Saccharoтyces, виды Nadsonia и др.) наблюдается так называемое почкующееся деление, при котором почка формируется на широком основании (рис. 18, 3). Почкующееся деление имеет морфологическое сходство с почкованием и делением. 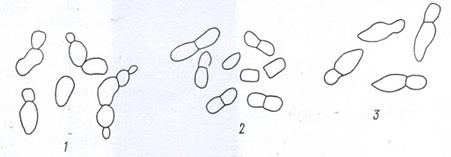
Рисунок 18. Способы вегетативного размножения дрожжей: 1 – почкование; 2 – деление; 3 – почкующееся деление
Бесполое и половое размножение связано с формированием специализированных репродуктивных структур. При половом размножении их появлению предшествует слияние клеток и последующее объединение ядер, при бесполом размножении предварительное слияние клеток и ядер не происходит.
Размножение бесполыми спорами - баллистоспорами - характерно для видов Sporoboloтyces, Sporidiobolus и др. Баллистоспоры возникают на заостренных выростах клеток - стеригмах (рис. 19). При созревании они с силой отбрасываются на некоторое расстояние. Баллистоспоровые дрожжи относятся к базидиомицетам. Истинный мицелий таких дрожжей может распадаться на 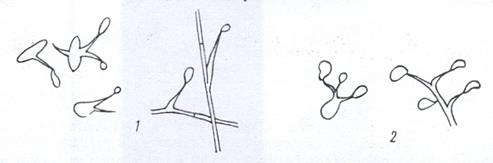
Рисунок 19. Баллистоспоры на отдельных клетках и гифах мицелия: 1 – Sporobolomyces; 2 - Sporidiobolus
отдельные клетки, называемые артроспорами.
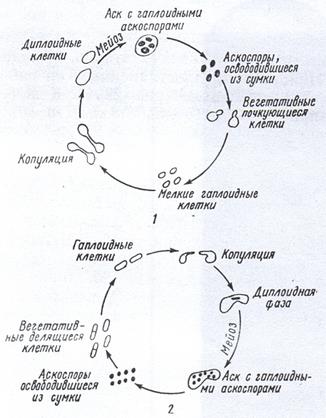
Рисунок 20. Жизненные циклы некоторых дрожжей: 1 – Saccharomyces cerevisiae; 2 – Schizosaccharomyces (Octosporomyces) octosporus
Половое размножение большинства дрожжей (аскомицетов) связано с образованием асков (сумок) и аскоспор. у одних (виды Saccharoтyces, Debaryoтyces и др.) образование асков происходит сразу после изо- или гетерогамной копуляции почек или вегетативных гаплоидных клеток в диплоидную. У других (виды Saccharoтyces, Schizosaccharoтyces, Saccharoтycodes и др.). Аски формируются из ранее возникших диплоидных клеток партеногенетически, т. е. без непосредственно предшествующей копуляции. В асках в результате мейоза образуются гаплоидные аскоспоры. В каждом аске бывает, как правило, от 2 до 8 спор. Иногда (виды Liротуces) их число достигает нескольких десятков. Аскоспоры многих дрожжей (виды Saccharoтyces, Schizosaccharoтyces) освобождаются из сумок и превращаются в вегетативные клет
Дата добавления: 2021-02-19; просмотров: 756;
Поиск по сайту
Узнать еще
- ОСНОВНЫЕ ТИПЫ И СВОЙСТВА НАПОЛЬНЫХ И БОРТОВЫХ СИСТЕМ ТЕХНИЧЕСКОГО ДИАГНОСТИРОВАНИЯ
- I. Гидрометаллургические методы
- I. Общая характеристика категории состояния как часть речи
- I. Погрешности механической обработки. Точность обработки. Методы их расчёта
- II. Лексико-грамматические разряды имен числительных. Их характеристика.
- II. Лексико-грамматические разряды местоимений. Их общая характеристика
- II. Методы исследования истории медицины.
- II. Основные положения

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
