Сенсорные системы человека и млекопитающих
10.4.1 Зрительная сенсорная система
Зрительная сенсорная система служит для восприятия и анализа световых раздражений. Через нее человек получает до 80-90 % всей информации о внешней среде. Глаз человека воспринимает световые лучи лишь в видимой части спектра — в диапазоне от 400 до 800 нм.
Зрительная сенсорная система состоит из следующих отделов:
1)периферический отдел — сложный вспомогательный орган — глаз, в котором находятся фоторецепторы и тела 1-х (биполярных) и 2-х (ганглиозных) нейронов;
2) проводниковый отдел — зрительный нерв (вторая пара черепно-мозговых нервов), представляющий собой волокна 2-ых нейронов и частично перекрещивающийся в хиазме, передает информацию третьим нейронам, часть которых расположена в переднем двухолмии среднего мозга, другая часть — в ядрах промежуточного мозга, так называемых наружных коленчатых телах;
3) корковый отдел — 4-е нейроны находятся в 17 поле затылочной области коры больших полушарий. Это образование представляет собой первичное (проекционное) поле или ядро анализатора, функцией которого является возникновение ощущений. Рядом с ним находится вторичное поле или периферия анализатора (18 и 19 поля), функция которого — опознание и осмысливание зрительных ощущений, что лежит в основе процесса восприятия. Дальнейшая обработка и взаимосвязь зрительной информации с информацией от других сенсорных систем происходит в ассоциативных задних третичных полях коры — нижнетеменных областях.
Глазное яблоко представляет собой шаровидную камеру диаметром около 2,5 см, содержащую светопроводящие среды — роговицу,влагу передней камеры, хрусталик и студнеобразную жидкость — стекловидное тело, назначение которых преломлять световые лучи и фокусировать их в области расположения рецепторов на сетчатке. Стенками камеры служат 3 оболочки. Наружная непрозрачная оболочка (склера) переходит спереди в прозрачную роговицу. Средняя сосудистая оболочка в передней части глаза образует ресничное тело и радужную оболочку, обусловливающую цвет глаз. В середине радужной оболочки (радужки) имеется отверстие — зрачок, регулирующий количество пропускаемых световых лучей. Диаметр зрачка регулируется зрачковым рефлексом, центр которого находится в среднем мозге. Внутренняя сетчатая оболочка (сетчатка) или содержит фоторецепторы глаза (палочки и колбочки) и служит для преобразования световой энергии в нервное возбуждение. Светопреломляющие среды глаза, преломляя световые лучи, обеспечивают четкое изображение на сетчатке. Основными преломляющими средами глаза человека являются роговица и хрусталик. Лучи, идущие из бесконечности через центр роговицы и хрусталика (т. е. через главную оптическую ось глаза) перпендикулярно к их поверхности, не испытывают преломления. Все остальные лучи преломляются и сходятся внутри камеры глаза в одной точке — фокусе. Приспособление глаза к четкому видению различно удаленных предметов (его фокусирование) называется аккомодацией. Этот процесс у человека осуществляется за счет изменения кривизны хрусталика. Ближняя точка ясного видения с возрастом отодвигается (от 7 см в 7-10 лет до 75 см в 60 лет и более), так как снижается эластичность хрусталика и ухудшается аккомодация. Возникает старческая дальнозоркость.
В норме длинник глаза соответствует преломляющей силе глаза. Однако у 35% людей имеются нарушения этого соответствия. В случае близорукости длинник глаза больше нормы и фокусировка лучей происходит перед сетчаткой, а изображение на сетчатке становится расплывчатым. В дальнозорком глазу, наоборот, длинник глаза меньше нормы и фокус располагается за сетчаткой. В результате изображение на сетчатке тоже расплывчато.
Рассмотрим механизм фоторецепции. Фоторецепторы глаза (палочки и колбочки) — это высокоспециализированные клетки, преобразующие световые раздражения в нервное возбуждение. Фоторецепция начинается в наружных сегментах этих клеток, где на специальных дисках, как на полочках, расположены молекулы зрительного пигмента (в палочках — родопсин, в колбочках — разновидности его аналога). Под действием света происходит ряд очень быстрых превращений и обесцвечивание зрительного пигмента.
В ответ на стимул эти рецепторы, в отличие от всех других рецепторов, формируют рецепторный потенциал в виде тормозных изменений на мембране клетки. Другими словами, на свету происходит гиперполяризация мембран рецепторних клеток, а в темноте — их деполяризация, т. е. стимулом для них является темнота, а не свет. При этом в соседних клетках происходят обратные изменения, что позволяет отделить светлые и темные точки пространства. Фотохимические реакции в наружных сегментах фоторецепторов вызывают изменения в мембранах остальной части рецепторной клетки, которые передаются биполярным клеткам (первым нейронам), а затем и ганглиозным клеткам (вторым нейронам), от которых нервные импульсы направляются в головной мозг. Часть ганглиозных клеток возбуждается на свету, часть — в темноте.
Палочки, рассеянные преимущественно по периферии сетчатки (их 130 млн), и колбочки, расположенные преимущественно в центральной части сетчатки (их 7 млн), различаются по своим функциям (рисунок 10.1-А). Палочки обладают более высокой чувствительностью, чем колбочки, и являются органами сумеречного зрения. Они воспринимают черно-белое (бесцветное) изображение. Колбочки представляют собой органы дневного зрения. Они обеспечивают цветное зрение. Существует 3 вида колбочек у человека: воспринимающие преимущественно красный, зеленый и сине-фиолетовый цвет. Разная их цветовая чувствительность определяется различиями в зрительном пигменте. Комбинации возбуждения этих приемников разных цветов дают ощущения всей гаммы цветовых оттенков, а равномерное возбуждение всех трех типов колбочек — ощущение белого цвета. При нарушении функции колбочек наступает цветовая слепота (дальтонизм), человек перестает различать цвета, в частности, красный и зеленый цвет. Это заболевание отмечается у 8% мужчин и у 0,5% женщин.
Важными функциональными характеристиками органа зрения являются острота и поле зрения
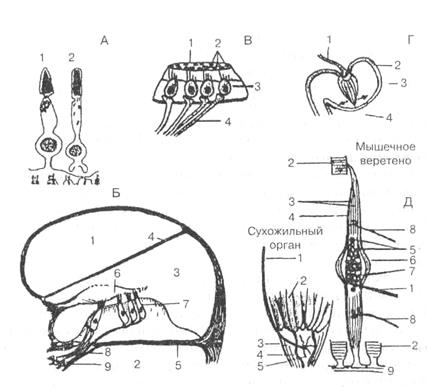
А: фоторецепторы. Колбочки (1) и палочки (2).
Б: слуховые рецепторы. 1 — вестибулярная лестница, 2 — барабанная
лестница, 3 — перепончатый канал улитки, 4 — вестибулярная мембрана,
5 — основная мембрана, 6 — покровная мембрана, 7— волосковые клетки,
8 — афферентные нервные волокна, 9 — нервные клетки спирального
ганглия (первые нейроны).
В и Г: вестибулярные рецепторы. В — отолитовый аппарат.
1 — отолитовая мембрана, 2 — отолиты (кристаллы углекислого кальция), 3 — волосковые рецепторные клетки,
4 — волокна вестибулярного нерва.
Г— полукружные каналы. I— волокно вестибулярного нерва, 2 — ампула, 3 — купула с волосковыми рецепторными клетками, 4 — полукружный канал. Стрелки показывают направление колебаний купулы при инерционных смещениях эндолимфы.
Д: проприорецепторы. Мышечное веретено. 1 — афферентное нервное
волокно, 2 — экстрафузальные мышечные волокна (перерезаны),
3 — внутриверетенные (интрафузальные) мышечные волокна,
4 — оболочка веретена, 5 — ядра, 6 — ядерная сумка, 7— чувствительные нервные окончания, 8 — эфферентные нервные гамма-волокна, 9 — сухожилие.
Сухожильный орган. I — афферентное нервное волокно, 2 — мышечные волокна, 3 — сухожилие, 4 — капсула, 5 — чувствительные нервные окончания.
Рисунок 10.1 – Рецепторы сенсорных систем
Остротой зрения называется способность различать отдельные объекты. Она измеряется минимальным углом, при котором две точки воспринимаются как раздельные, — примерно 0.5 угловой минуты. В центре сетчатки колбочки имеют более мелкие размеры и расположены гораздо плотнее, поэтому способность к пространственному различению здесь в 4-5 раз выше, чем на периферии сетчатки. Следовательно, центральное зрение отличается более высокой остротой зрения, чем периферическое зрение.
Для детального разглядывания предметов человек поворотом головы и глаз перемещает их изображение в центр сетчатки.
Острота зрения зависит не только от густоты рецепторов, но и от четкости изображения на сетчатке, т. е, от преломляющих свойств глаза, от степени аккомодации, от величины зрачка. В водной среде преломляющая сила роговицы снижается, так как ее коэффициент преломления близок к коэффициенту воды. В результате под водой острота зрения уменьшается в 200 раз.
Полем зрения называется часть пространства, видимая при неподвижном положении глаза. Для черно-белых сигналов поле зрения обычно ограничено строением костей черепа и положением в глазницах глазных яблок. Для цветных раздражителей поле зрения меньше, так как воспринимающие их колбочки находятся в центральной части сетчатки. Наименьшее поле зрения отмечается для зеленого цвета. При утомлении поле зрения уменьшается.
Человек обладает бинокулярным зрением, т.е. зрением двумя глазами. Такое зрение имеет преимущество перед монокулярным зрением (одним глазом) в восприятии глубины пространства, особенно на близких расстояниях (менее 100 м).
Четкость такого восприятия (глазомер) обеспечивается хорошей координацией движения обоих глаз, которые должны точно наводиться на рассматриваемый объект. В этом случае его изображение попадает на идентичные точки сетчатки (одинаково удаленные от центра сетчатки) и человек видит одно изображение.
Четкий поворот глазных яблок зависит от работы наружных мышц глаза — его глазодвигательного аппарата (четырех прямых и двух косых мышц), другими словами, от мышечного баланса глаза. Однако идеальный мышечный баланс глаза или ортофория имеется лишь у 40%людей. Его нарушение возможно в результате утомления, действия алкоголя и пр., а также как следствие дисбаланса мышц, что приводит к нечеткости и раздвоению изображения (гетерофория).
При небольших нарушениях сбалансированности мышечных усилий наблюдается небольшое скрытое (или физиологическое) косоглазие, которое в бодром состоянии человек компенсирует волевой регуляцией, а при значительных — явное косоглазие.
Глазодвигательный аппарат имеет большое значение в восприятии скорости движения, которую человек оценивает либо по скорости перемещения изображения по сетчатке неподвижного глаза, либо по скорости движения наружных мышц глаза при следящих движениях глаза.
Изображение, которое видит человек двумя глазами, прежде всего, определяется его ведущим глазом. Ведущий глаз обладает более высокой остротой зрения, мгновенным и особенно ярким восприятием цвета, более обширным полем зрения, лучшим ощущением глубины пространства.
При прицеливании воспринимается лишь то, что входит в поле зрения этого глаза. В целом, восприятие объекта в большей мере обеспечивается ведущим глазом, а восприятие окружающего фона — не ведущим глазом.
10.4.2 Слуховая сенсорная система (рисунок 10.1 – Б).
Слуховая сенсорная система служит для восприятия и анализа звуковых колебаний внешней среды. Она приобретает у человека особо большое значение в связи с развитием речевого общения между людьми. Деятельность слуховой сенсорной системы имеет также значение для оценки временных интервалов — темпа и ритма движений.
Слуховая сенсорная система состоит из следующих разделов:
1)периферический отдел, который представляет собой сложный специализированный орган, состоящий из наружного,среднего и внутреннего уха;
2) проводниковый отдел — первый нейрон проводникового отдела, находящийся в спиральном узле улитки, получает возбуждение от рецепторов внутреннего уха, отсюда информация поступает по его волокнам, т. е. по слуховому нерву (входящему в 8 пар черепно-мозговых нервов), ко второму нейрону в продолговатом мозге, а после перекреста часть волокон идет к третьему нейрону в заднем двухолмии среднего мозга, а часть к ядрам промежуточного мозга — внутреннему коленчатому телу;
3) корковый отдел — представлен четвертым нейроном, который находится в первичном (проекционном) слуховом поле в височной области коры больших полушарий и обеспечивает возникновение ощущения, а более сложная обработка звуковой информации происходит в расположенном рядом вторичном слуховом поле, отвечающем за формирование восприятия и опознание информации. Полученные сведения поступают в третичное поле нижнетеменной зоны, где интегрируются с другими формами информации.
Рассмотрим функции наружного, среднего и внутреннего уха.
Наружное ухо является звукоулавливающим аппаратом. Звуковые колебания улавливаются ушными раковинами (у животных они могут поворачиваться к источнику звука) и передаются по наружному слуховому проходу к барабанной перепонке, которая отделяет наружное ухо от среднего. Улавливание звука и весь процесс слушания двумя ушами (бинауральный слух) имеет значение для определения направления звука. Звуковые колебания, идущие сбоку, доходят до ближайшего уха на несколько десятитысячных долей секунды (0,0006 с) раньше, чем до другого уха. Этой ничтожной разницы во времени прихода звука к обоим ушам достаточно, чтобы определить его направление.
Среднее ухо является звукопроводящим аппаратом. Оно представляет собой воздушную полость, которая через слуховую (евстахиеву) трубу соединяется с полостью носоглотки. Колебания от барабанной перепонки через среднее ухо передают соединенные друг с другом 3 слуховые косточки — молоточек, наковальня и стремечко, а последнее через перепонку овального окна передает эти колебания жидкости, находящейся во внутреннем ухе, — перилимфе. Благодаря слуховым косточкам амплитуда колебаний уменьшается, а сила их увеличивается, что позволяет приводить в движение столб жидкости во внутреннем ухе. При сильных звуках специальные мышцы уменьшают подвижность барабанной перепонки и слуховых косточек, адаптируя слуховой аппарат к таким изменениям раздражителя и предохраняя внутреннее ухо от разрушения. Благодаря соединению через слуховую трубу воздушной полости среднего уха с полостью носоглотки возникает возможность выравнивания давления по обе стороны барабанной перепонки, что предотвращает ее разрыв при значительных изменениях давления во внешней среде — при погружениях под воду, подъемах на высоту, выстрелах и пр. Это барофункция уха.
Внутреннее ухо является звуковоспринимающим аппаратом. Оно расположено в пирамиде височной кости и содержит улитку, которая у человека образует 2,5 спиральных витка. Улитковый канал разделен двумя перегородками основной мембраной и вестибулярной мембраной на 3 узких хода: верхний (вестибулярная лестница), средний (перепончатый канал) и нижний (барабанная лестница). На вершине улитки имеется отверстие, соединяющее верхний и нижний каналы в единый, идущий от овального окна к вершине улитки и далее к круглому окну. Полость его заполнена жидкостью — перилимфой, а полость среднего перепончатого канала заполнена жидкостью иного состава — эндолимфой. В среднем канале расположен звуковоспринимаюший аппарат — Кортиев орган, в котором находятся механорецепторы звуковых колебаний — волосковые клетки.
Рассмотрим физиологический механизм восприятия звука. Восприятие звука основано на двух процессах, происходящих в улитке: разделение звуков различной частоты по месту их наибольшего воздействия на основную мембрану улитки; преобразование рецепторными клетками механических колебаний в нервное возбуждение.
Звуковые колебания, поступающие во внутреннее ухо через овальное окно, передаются перилимфе, а колебания этой жидкости приводят к смещениям основной мембраны. От высоты звука зависит высота столба колеблющейся жидкости и соответственно место наибольшего смещения основной мембраны: звуки высокой частоты дают наибольший эффект на начале основной мембраны, а звуки низких частот доходят до вершины улитки. Таким образом, при различных по частоте звуках возбуждаются разные волосковые клетки и разные нервные волокна, т. е. осуществляется пространственный код. Увеличение силы звука приводит к увеличению числа возбужденных волосковых клеток и нервных волокон, что позволяет различать интенсивность звуковых колебаний.
Волоски рецепторных клеток погружены в покровную мембрану. При колебаниях основной мембраны начинают смещаться находящиеся на ней волосковые клетки и их волоски механически раздражаются покровной мембраной. В результате в волосковых рецепторах возникает процесс возбуждения, который по афферентным волокнам направляется к нейронам спирального узла улитки и далее в ЦНС.
Различают костную и воздушную проводимость звука. В обычных условиях у человека преобладает воздушная проводимость — проведение звуковых колебаний через наружное и среднее ухо к рецепторам внутреннего уха. В случае костной проводимости звуковые колебания передаются через кости черепа непосредственно улитке (например, при нырянии, подводном плавании).
Человек обычно воспринимает звуки с частотой от 15 до 20000 Гц (в диапазоне 10-11 октав). У детей верхний предел достигает 22000 Гц, с возрастом он понижается. Наиболее высокая чувствительность обнаружена в области частот от 1000 до 3000 Гц. Эта область соответствует наиболее часто встречающимся частотам человеческой речи и музыки.
10.4.3 Вестибулярная сенсорная система (рисунок 10.1 – В,Г)
Вестибулярная сенсорная система служит для анализа положения и движения тела в пространстве. Это одна из древнейших сенсорных систем, развившаяся в условиях действия силы тяжести на земле. Импульсы вестибулярного аппарата используются в организме для поддержания равновесия тела, для регуляции и сохранения позы, для пространственной организации движений человека.
Вестибулярная сенсорная система состоит из следующих отделов:
1) периферический отдел: включает два образования,содержащие механорецепторы вестибулярной системы — преддверие(мешочек и маточка) и полукружные каналы;
2) проводниковый отдел: начинается от рецепторов волокнами первого нейрона вестибулярного узла, расположенного в височной кости; другие отростки этих нейронов образуют вестибулярный нерв и вместе со слуховым нервом в составе 8-ой пары черепно-мозговых нервов входят в продолговатый мозг; в вестибулярных ядрах продолговатого мозга находятся вторые нейроны, импульсы от которых поступают к третьим нейронам (в таламус промежуточного мозга);
3) корковый отдел представляют четвертые нейроны, часть которых представлена в проекционном (первичном) поле вестибулярной системы в височной области коры, а другая часть — находится в непосредственной близости к пирамидным нейронам моторной области коры и в постцентральной извилине. Точная локализация коркового отдела вестибулярной сенсорной системы у человека в настоящее время не установлена.
Рассмотрим особенности функционирования вестибулярного аппарата. Периферический отдел вестибулярной сенсорной системы находится во внутреннем ухе. Каналы и полости в височной кости образуют костный лабиринт вестибулярного аппарата, который частично заполнен перепончатым лабиринтом. Между костным и перепончатым лабиринтами находится жидкость — перилимфа, а внутри перепончатого лабиринта — эндолимфа.
Аппарат преддверия предназначен для анализа действия силы тяжести при изменениях положения тела в пространстве и ускорений прямолинейного движения. Перепончатый лабиринт преддверия разделен на 2 полости — мешочек и маточку, содержащих отолитовые приборы. Механорецепторы отолитовых приборов представляют собой волосковые клетки. Они склеены студнеобразной массой, образующей поверх волосков отолитовую мембрану, в которой находятся кристаллы углекислого кальция — отолиты. В маточке отолитовая мембрана расположена в горизонтальной плоскости, а в мешочке она согнута и находится во фронтальной и сагиттальной плоскостях. При изменении положения головы и тела, а также при вертикальных или горизонтальных ускорениях отолитовые мембраны свободно перемещаются под действием силы тяжести во всех трех плоскостях, натягивая, сжимая или сгибая при этом волоски механорецепторов. Чем больше деформация волосков, тем выше частота афферентных импульсов в волокнах вестибулярного нерва.
Аппарат полукружных каналов служит для анализа действия центробежной силы при вращательных движениях. Адекватным его раздражителем является угловое ускорение. Три дуги полукружных каналов распложены в трех взаимно перпендикулярных плоскостях. Передняя дуга находится во фронтальной плоскости, боковая — в горизонтальной плоскости, задняя — в сагиттальной плоскости. В одном из концов каждого канала имеется расширение — ампула. Находящиеся в ней волоски чувствительных клеток склеены в гребешок — ампулярную купулу. Она представляет собой маятник, который может отклоняться в результате разности давления эндолимфы на противоположные поверхности купулы. При вращательных движениях в результате инерции эндолимфа отстает от движения костной части и оказывает давление на одну из поверхностей купулы. Отклонение купулы изгибает волоски рецепторных клеток и вызывает появление нервных импульсов в вестибулярном нерве. Наибольшие изменения в положении купулы происходят в том полукружном канале, положение которого соответствует плоскости вращения.
В настоящее время показано, что вращения или наклоны в одну сторону увеличивают афферентную импульсацию, а в другую сторону— уменьшают ее. Это позволяет различать направление прямолинейного или вращательного движения.
Вестибулярная система оказывает определенные влияния на другие функции организма. Она связана со многими центрами спинного и головного мозга и вызывает ряд вестибуло-соматических и вестибуло-вегетативных рефлексов.
Вестибулярные раздражения вызывают установочные рефлексы изменения тонуса мышц, лифтные рефлексы, а также особые движения глаз, направленные на сохранение изображения на сетчатке, — нистагм (движения глазных яблок со скоростью вращения, но в противоположном направлении, затем быстрое возвращение к исходной позиции и новое противоположное вращение).
Помимо основной анализаторной функции, важной для управления позой и движениями человека, вестибулярная сенсорная система оказывает разнообразные побочные влияния на многие функции организма, которые возникают в результате иррадиации возбуждения на другие нервные центры при низкой устойчивости вестибулярного аппарата. Его раздражение приводит к снижению возбудимости зрительной и кожной сенсорных систем, ухудшению точности движений. Вестибулярные раздражения приводят к нарушениям координации движений и походки, изменениям частоты сердцебиения и артериального давления, увеличению времени двигательной реакции и снижению частоты движений, ухудшению чувства времени, изменению психических функций — внимания, оперативного мышления, кратковременной памяти, эмоциональных проявлений, В тяжелых случаях возникают головокружения, тошнота, рвота. Повышение устойчивости вестибулярной системы достигается в большей мере активными вращениями человека, чем пассивными.
В условиях невесомости (когда у человека выключены вестибулярные влияния) возникает утрата представления о направлении гравитационной вертикали и пространственном положении тела. Теряются навыки ходьбы, бега. Ухудшается состояние нервной системы, возникает повышенная раздражительность, нестабильность настроения.
10.4.4 Двигательная сенсорная система (рисунок 10.1 – Д)
Двигательная сенсорная система служит для анализа состояния двигательного аппарата — его движения и положения. Информация о степени сокращения скелетных мышц, натяжении сухожилий, изменении суставных углов необходима для регуляции двигательных актов и поз.
Двигательная сенсорная система состоит из следующих 3-х отделов:
1) периферический отдел, представленный проприорецепторами, расположенными в мышцах, сухожилиях и суставных сумках;
2) проводниковый отдел, который начинается биполярными клетками (первыми нейронами), тела которых расположены вне ЦНС — в спинномозговых узлах. Один их отросток связан с рецепторами, другой входит в спинной мозг и передает проприоцептивные импульсы ко вторым нейронам в продолговатый мозг (часть путей от проприорецепторов направляется в кору мозжечка), а далее к третьим нейронам — релейным ядрам таламуса (в промежуточный мозг);
3) корковый отдел находится в передней центральной извилине коры больших полушарий.
Рассмотрим функции проприорецепторов. К проприорецепторам относятся мышечные веретена, сухожильные органы (или органы Гольджи) и суставные рецепторы (рецепторы суставной капсулы и суставных связок). Все эти рецепторы представляют собой механорецепторы, специфическим раздражителем которых является их растяжение.
Мышечные веретена прикрепляются к мышечным волокнам параллельно — один конец к сухожилию, а другой — к волокну. Каждое веретено покрыто капсулой, образованной несколькими слоями клеток, которая в центральной части расширяется и образует ядерную сумку. Внутри веретена содержится несколько (от 2 до 14) тонких внутри веретенных или так называемых интрафузальных мышечных волокон. Эти волокна в 2-3 раза тоньше обычных волокон скелетных мышц (экстрафузальных).
Интрафузалъные волокна подразделяются на два типа: длинные, толстые, с ядрами в ядерной сумке, которые связаны с наиболее толстыми и быстропроводящими афферентными нервными волокнами — они информируют о динамическом компоненте движения (скорости изменения длины мышцы); короткие, тонкие, с ядрами, вытянутыми в цепочку, информирующие о статическом компоненте (удерживаемой в данный момент длине мышцы).
Окончания афферентных нервных волокон намотаны на интрафузальные волокна рецептора. При растяжении скелетной мышцы происходит растяжение и мышечных рецепторов, которое деформирует окончания нервных волокон и вызывает появление в них нервных импульсов. Частота проприоцептивной импульсации возрастает с увеличением растяжения мышцы, а также при увеличении скорости ее растяжения. Тем самым нервные центры информируются о скорости растяжения мышцы и ее длине. Вследствие малой адаптации импульсация от мышечных веретен продолжается в течение всего периода поддержания растянутого состояния, что обеспечивает постоянную осведомленность центров о длине мышцы. Чем более тонкие и координированные движения осуществляют мышцы, тем больше в них мышечных веретен: у человека в глубоких мышцах шеи, связывающих позвоночник с головой, среднее их число составляет 63, а в мышцах бедра и таза—менее 5 веретенна 1 г массы мышцы.
ЦНС может тонко регулировать чувствительность проприорецепторов. Разряды мелких гамма-мотонейронов спинного мозга вызывают сокращение интрафузальных мышечных волокон по обе стороны от ядерной сумки веретена. В результате средняя несократимая часть мышечного веретена растягивается, и деформация отходящего отсюда нервного волокна вызывает повышение
его возбудимости. При той же длине скелетной мышцы в нервные центры при этом будет поступать большее число афферентных импульсов. Это позволяет, во-первых, выделять проприоцептивную им пульсацию на фоне другой афферентной информации и, во-вторых, увеличивать точность анализа состояния мышц. Повышение чувствительности веретен происходит во время движения и даже в предстартовом состоянии. Это объясняется тем, что в силу низкой возбудимости гамма-мотонейронов их активность в состоянии покоя выражена слабо, а при произвольных движениях и вестибулярных реакциях она активируется. Чувствительность проприорецепторов повышается также при умеренных раздражениях симпатических волокон и выделении небольших доз адреналина.
Сухожильные органы расположены в месте перехода мышечных волокон в сухожилия. Сухожильные рецепторы (окончания нервных волокон) оплетают тонкие сухожильные волокна, окруженные капсулой. В результате последовательного крепления сухожильных органов к мышечным волокнам (а в ряде случаев — к мышечным веретенам), растяжение сухожильных механорецепторов происходит при напряжении мышц. Таким образом, в отличие от мышечных веретен, сухожильные рецепторы информируют нервные центры о степени напряжения мышц и скорости его развития.
Суставные рецепторы информируют о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы представляют собой свободные нервные окончания или окончания, заключенные в специальную капсулу. Одни суставные рецепторы посылают информацию о величине суставного угла, т. е. о положении сустава. Их импульсация продолжается в течение всего периода сохранения данного угла. Она тем большей частоты, чем больше сдвиг угла. Другие суставные рецепторы возбуждаются только в момент движения в суставе, т. е. посылают информацию о скорости движения. Частота их импульсации возрастает с увеличением скорости изменения суставного угла.
Сигналы, идущие от рецепторов мышечных веретен, сухожильных органов, суставных сумок и тактильных рецепторов кожи, называют кинестетическими, т. е. информирующими о движении тела. Их участие в произвольной регуляции движений различно. Сигналы от суставных рецепторов вызывают заметную реакцию в коре больших полушарий и хорошо осознаются. Благодаря им человек лучше воспринимает различия при движениях в суставах, чем различия в степени напряжения мышц при статических положениях или поддержании веса. Сигналы же от других проприорецепторов, поступающие преимущественно в мозжечок, обеспечивают бессознательную регуляцию, подсознательный контроль движений и поз.
10.4.5 Сенсорные системы кожи, внутренних органов, вкуса и обоняния
В коже и внутренних органах имеются разнообразные рецепторы, реагирующие на физические и химические раздражители.
Рассмотрим особенности кожной рецепции.
В коже представлена тактильная, температурная и болевая рецепция. На 1 см 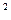 кожи, в среднем, приходится 12-13 холодовых точек, 1 -2 тепловых, 25 тактильных и около 100 болевых.
кожи, в среднем, приходится 12-13 холодовых точек, 1 -2 тепловых, 25 тактильных и около 100 болевых.
Тактильная сенсорная система предназначена для анализа давления и прикосновения. Ее рецепторы представляют собой свободные нервные окончания и сложные образования (тельца Мейснера, тельца Паччини), в которых нервные окончания заключены в специальную капсулу.
Они находятся в верхних и нижних слоях кожи, в кожных сосудах, в основаниях волос. Особенно их много на пальцах рук и ног, ладонях, подошвах, губах. Это механорецепторы, реагирующие на растяжение, давление и вибрацию. Наиболее чувствительным рецептором является тельце Паччини, которое вызывает ощущение прикосновения при смещении капсулы лишь на 0.0001 мм. Чем больше размеры тельца Паччини, тем более толстые и быстропроводяшие афферентные нервы отходят от него. Они проводят кратковременные залпы (длительностью 0.005 с), информирующие о начале и окончании действия механического раздражителя. Путь тактильной информации следующий: рецептор — 1 -и нейрон в спинномозговых узлах —2-й нейрон в спинном или продолговатом мозге — 3-й нейрон в промежуточном мозге (таламус) — 4-й нейрон в задней центральной извилине коры больших полушарий (первичная соматосенсорная зона).
Температурная рецепция осуществляется Холодовыми рецепторами (колбы Краузе) и тепловыми (тельца Руффини, Гольджи-Маццони). При температуре кожи 31-37°С эти рецепторы почти неактивны. Ниже этой границы холодовые рецепторы активизируются пропорционально падению температуры, затем их активность падает и совсем прекращается при +12 С. При температуре выше 37°С активизируются тепловые рецепторы, достигают максимальной активности при +43°С, а затем они резко прекращают ответы.
Болевая рецепция, как считает большинство специалистов, не имеет специальных воспринимающих образований. Болевые раздражения воспринимаются свободными нервными окончаниями, а также возникают при сильных температурных и механических раздражениях в соответствующих термо- и механорецепторах.
Температурные и болевые раздражения передаются в спинной мозг, оттуда в промежуточный мозг и в соматосенсорную область коры.
Рассмотрим устройство висцероцептивной (интероцептивной) сенсорной системы.
Во внутренних органах имеется множество рецепторов, воспринимающих давление — барорецепторы сосудов, кишечного тракта и др., изменения химизма внутренней среды — хеморецепторы, ее температуры — терморецепторы, осмотического давления, болевые раздражения. С их помощью безусловнорефлекторным путем регулируется постоянство различных констант внутренней среды (поддержание гомеостаза), ЦНС информируется об изменениях во внутренних органах. Информация от интерорецепторов через блуждающий, чревный и тазовый нервы поступает в промежуточный мозг и далее в , лобные и другие области коры головного мозга. Деятельность этой системы практически не осознается, она мало локализована, однако при сильных раздражениях она хорошо ощущается. Она участвует в формировании сложных ощущений — жажды, голода и др.
Обонятельная и вкусовая сенсорные системы относятся к древнейшим системам. Они предназначены для восприятия и анализа химических раздражений, поступающих из внешней среды. Хеморецепторы обоняния находятся в обонятельном эпителии верхних носовых ходов.
Это — волосковые биполярные клетки, передающие информацию через решетчатую кость черепа к клеткам обонятельной луковицы мозга и далее через обонятельный тракт к обонятельным зонам коры (крючок морского коня, извилина гиппокампа и другие). Различные рецепторы избирательно реагируют на разные молекулы пахучих веществ, возбуждаясь лишь теми молекулами, которые являются зеркальной копией поверхности рецептора. Они воспринимают эфирный, камфарный, мятный, мускусный и др. запахи, причем к некоторым веществам чувствительность необычайно высока.
Хеморецепторы вкуса представляют собой вкусовые луковицы, расположенные в эпителии языка, задней стенке глотки и мягкого неба. У детей их количество больше, а с возрастом — убывает. Микроворсинки рецепторных клеток выступают из луковицы на поверхность языка и реагируют на растворенные в воде вещества. Их сигналы поступают через волокна лицевого и языкоглоточного нервов (продолговатый мозг) в таламус и далее (в соматосенсорную область коры). Рецепторы разных частей языка воспринимают четыре основных вкуса: горького (задняя часть языка), кислого (края языка), сладкого (передняя часть языка) и соленого (передняя часть и края языка). Между вкусовыми ощущениями и химическим строением вещества отсутствует строгое соответствие, так как вкусовые ощущения могут изменяться при заболевании, беременности, условно-рефлекторных воздействиях, изменениях аппетита. В формировании вкусовых ощущений участвуют обоняние, тактильная, болевая и температурная чувствительность. Информация вкусовой сенсорной системы используется для организации пищевого поведения, связанного с добыванием, выбором, предпочтением или отверганием пищи, формированием чувства голода, сытости.
Как же происходит переработка сенсорной информации и каково ее значение? Сенсорная информация передается от рецепторов в высшие отделы мозга по двум основным путям нервной системы — специфическим и неспецифическим. Специфические проводящие пути составляют один из трех основных функциональных блоков мозга — блок приема, переработки и хранения информации. Это классические афферентные пути зрительной, слуховой, двигательной и др. сенсорных систем. В обработке этой информации участвует и неспецифическая система мозга, не имеющая прямых связей с периферическими рецепторами, но получающая импульсы по коллатералям от всех восходящих специфических систем и обеспечивающая их широкое взаимодействие.
Далее осуществляетс
Дата добавления: 2016-09-06; просмотров: 2819;
Поиск по сайту
Узнать еще
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- I.2. Антигены системы АВ0. Генетика. Структура
- I.2.1 ПЕРВЫЙ ЗАКОН НЬЮТОНА. ИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА.
- I.2.1 ПОЛНАЯ И ВНУТРЕННЯЯ ЭНЕРГИЯ СИСТЕМЫ. ТЕПЛОТА И РАБОТА
- I.3. Антитела системы АВ0
- I.5.4 НЕИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА. СИЛЫ ИНЕРЦИИ
- II. ЭЛЕКТРИЧЕСКИЙ ДИПОЛЬ. ДИПОЛЬНЫЙ МОМЕНТ СИСТЕМЫ ЭЛЕКТРИЧЕСКИХ ЗАРЯДОВ
- III. Избирательные системы.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
