Симпатический и парасимпатический отделы автономной нервной системы
В настоящее время всю автономную нервную систему подразделяют на симпатическую, парасимпатическую и метасимпатическую. Метасимпатическая – это нервная система, которая расположена внутри органа. Ее образует интрамуральный ганглий с системой нервных волокон.
Высшим отделом симпатической и парасимпатической иннервации является гипоталамус.
Медиаторами автономной нервной системы являются те же, что и в ЦНС: адреналин, норадреналин, ацетилхолин, вещество Р, гамма-амино-
масляная кислота, гистамин, серотонин и др.
Существуют определенные различия в морфофункциональной организации симпатического и парасимпатического отделов автономной иннервации.
1. Отличие проявляется в локализации центральных отделов симпатической и парасимпатической иннервации: центры симпатической нервной системы локализуются в основном в боковых рогах тораколюмбального отдела спинного мозга, а центры парасимпатической иннервации находятся в стволе мозга и в крестцовом отделе спинного мозга. Так, преганглионарные волокна парасимпатической нервной системы, снабжающие глазные мышцы и железы головы, покидают ствол мозга в составе трех пар черепно-мозговых нервов – III (глазодвигательный), VII (лицевой) и IX (языкоглоточный). К органам грудной и брюшной полости подходят в составе Х пары (блуждающий нерв) преганглионарные волокна, а к органам таза в составе тазовых нервов – парасимпатические волокна крестцового отдела спинного мозга.
2. Отличие проявляется также в типе ганглиев. Для симпатической нервной системы характерны паравертебральные и превертебральные ганглии, для парасимпатической – превертебральные и интрамуральные ганглии.
3. Различаются эти отделы и по длине пре- и постганглионарных волокон. Так, для симпатической нервной системы характерным является большей частью меньшая длина преганглионарных волокон, чем постганглионарных. Для парасимпатической нервной системы, наоборот, длина преганглионарных волокон может намного превышать длину постганглионарных.
4. Различие существует в медиаторах, которые осуществляют передачу в синапсах между иннервируемым органом и постганглионарным волокном. Медиатор в вегетативных ганглиях является единым для симпатического и парасимпатического отделов – это ацетилхолин, реализующий свое влияние через Н-холинорецепторы. В синапсах между постганглионарным волокном и эффекторным органом эти медиаторы разные. Для симпатической иннервации медиатором является адреналин и норадреналин, который связывается с α- и β-адренорецепторами на постсинаптической мембране эффекторного органа. Для парасимпатической иннервации это ацетилхолин, который реализует свое влияние через М-холинорецеп-
торы на постсинаптической мебране эффекторного органа.
5. Физиологические эффекты при возбуждении симпатических или парасимпатических волокон у органов, имеющих двойную иннервацию, являются противоположными. Например, при возбуждении волокон блуждающего нерва, иннервирующих сердце, наблюдаются четыре отрицательных эффекта: инотропный (уменьшение силы сокращения), хронотропный (уменьшение частоты сокращения), батмотропный (уменьшение возбудимости) и дромотропный (уменьшение проводимости). При раздражении симпатических волокон, иннервирующих сердце, наблюдаются положительные эффекты. Блуждающий нерв для желудка является главным и секреторным, и моторным: активация его приводит к увеличению моторной и секреторной функции желудка. Активация симпатической нервной системы оказывает ингибирующий эффект на секреторную и моторную функцию желудка.
Следует заметить, что эффекты, вызываемые раздражением симпатических волокон, зависят от рецепторов на постсинаптической мембране эффекторного органа. Так, при действии адреналина на α-рецепторы происходит сужение артерий и артериол скелетных мышц, а при действии адреналина на β-рецепторы в сердечной мышце сосуды в сердце расширяются. Механизм работы адренергических синапсов представлен на схемах, приведенных ниже (рис. 2.13-2.16).

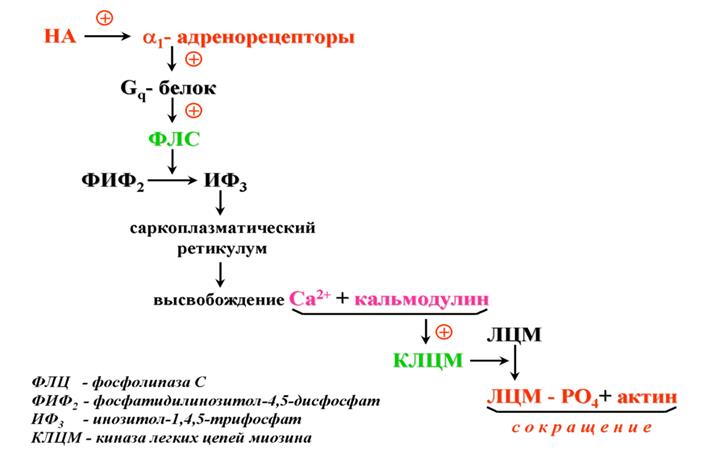
Основные эффекты:
1) расширение зрачков (сокращение радиальной мышцы радужки);
2) сужение кровеносных сосудов.
| |
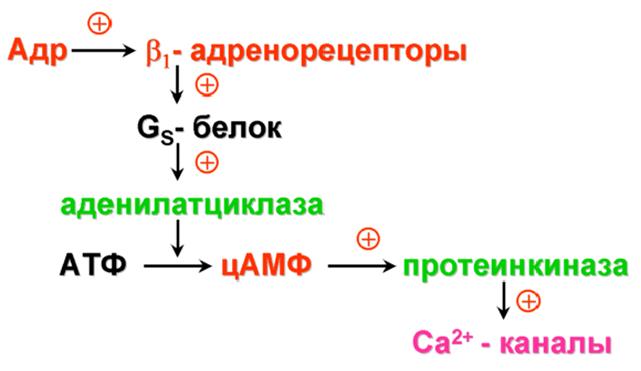
Основные эффекты:
· стимуляция деятельности сердца:
1) повышение силы сокращений;
2) повышение частоты сокращений;
3) облегчение атриовентрикулярной проводимости;
4) повышение автоматизма волокон проводящей системы.
| |
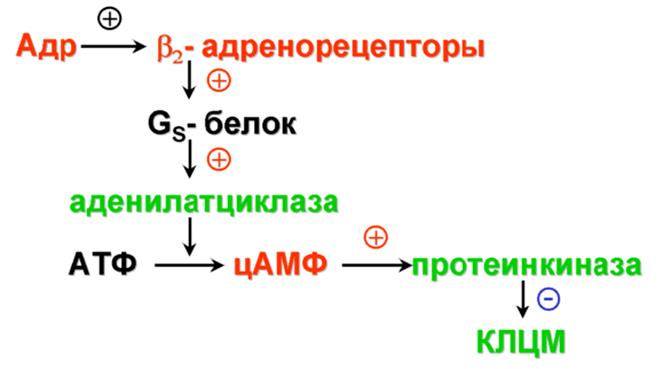
Основные эффекты:
1) расширение кровеносных сосудов;
2) расслабление гладких мышц бронхов;
3) снижение тонуса и ритмической сократительной активности миометрия;
4) гликогенолиз.
| |
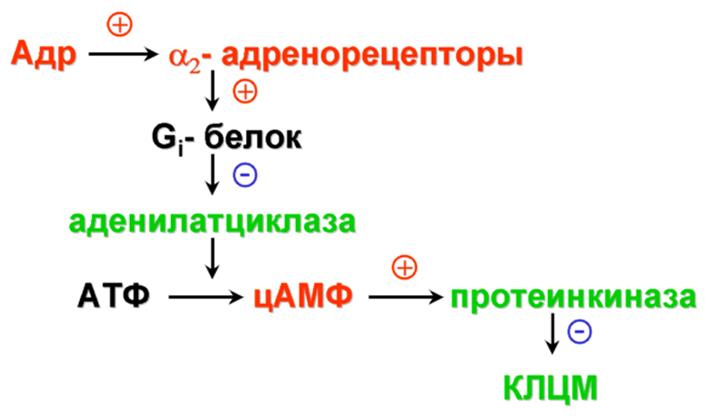
Основной эффект:
сужение кровеносных сосудов
Рис. 2.16. Механизм активации α2-адренорецепторов
| |
ЗАКЛЮЧЕНИЕ
Центральная нервная система, состоящая из отдельных нейронов, связанных синаптическими контактами, представляет собой единое образование. Благодаря морфофункциональным особенностям нейронов, процессам торможения, различным медиаторным системам, по принципу доминанты в ЦНС создаются направленные потоки возбуждения, которые позволяют целостно и одновременно дифференцированно вовлекать мозг в любую из форм его активности.
В работах И.П. Павлова и его учеников была высказана идея о динамической локализации функций. Эта идея получила дальнейшее развитие в работах физиологов, морфологов и клиницистов. В результате полученных фактов были разработаны принципы структурной и функциональной организации мозга и сформулирован эволюционный закон кортикализации функций. К настоящему времени убедительно доказано, что высшие функции мозга являются результатом системной деятельности мозга и в связи с этим поражение любого из отделов мозга может привести к распаду всей системы; таким образом, нарушение или выпадение функций не может непосредственно говорить о ее локализации.
Динамический характер деятельности системы и межсистемной интеграции определяется особенностями поступающей афферентной импульсации, специфической реакцией организма и его внутренней активностью. Динамичность этих взаимоотношений имеет свои особенности на поведенческом, нейронном, синаптическом и молекулярном уровнях интеграции. С позиций системной организации функций в деятельности мозга выделяют различные функциональные системы и подсистемы (Анохин П.К., Лурия А.Р., Соколов Е.М., Адрианов О.С., Батуев А.С., Судаков К.В. и др.). Классический вариант интегративной деятельности мозга может быть представлен в виде взаимодействия трех основных функциональных блоков:
1) блока приема и переработки информации – сенсорные системы (анализаторы);
2) блока модуляции, активации нервной системы (лимбико-ретику-
лярные системы мозга);
3) блока программирования, запуска и контроля поведенческих актов – моторные системы (двигательный анализатор).
1) Блок приема и контроля осуществляет процесс рецепции: в рецепторах сенсорных систем происходит трансформация энергии различных раздражителей в рецепторный и генераторный потенциалы; кодирования – на аксоне сенсорного нейрона происходит генерация потенциала действия и передача его по специфическим и неспецифическим путям в различные структуры ЦНС; перекодирования, анализа и синтеза. Этот процесс осуществляется в проекционных и ассоциативных областях коры, задействуются детекторные и гностические нейроны.
2) Модулирующие системы мозга регулируют тонус коры и подкорковых образований, оптимизируют уровень бодрствования в отношении выполняемой деятельности и обусловливают адекватный выбор поведения в соответствии с актуализируемой потребностью. Только в условиях оптимального бодрствования человек может наилучшим образом принимать и перерабатывать информацию, вызывать в памяти нужные избирательные системы связей, программировать деятельность, осуществлять контроль над ней. Аппаратом, выполняющим функцию регулятора уровня бодрствования, а также осуществляющим избирательную модуляцию и актуализацию приоритета той или иной функции, является модулирующая система мозга. Ее часто называют лимбико-ретикулярный комплекс или восходящая активирующая система. К нервным образованиям этого аппарата относятся лимбическая и неспецифическая системы мозга с активирующими и инактивирующими структурами. Среди активирующих мозговых структур прежде всего выделяют ретикулярную формацию (РФ) среднего мозга, задний гипоталамус, синее пятно в нижних отделах ствола мозга. К инактивирующим структурам относят преоптическую область гипоталамуса, ядра шва в стволе мозга, фронтальную кору.
Волокна ретикулярной формации, направляясь вверх, образуют модулирующие «входы» (аксо-дендритные синапсы) в вышерасположенных мозговых образованиях, включая старую и новую кору. От старой и новой коры берут начало нисходящие волокна, которые идут в обратном направлении к структурам гипоталамуса, среднего мозга и к более низким уровням мозгового ствола. Через нисходящие системы связей все нижележащие образования оказываются под управлением и контролем тех программ, которые возникают в коре головного мозга и для выполнения которых требуется модуляция активности и модификация состояния бодрствования. Таким образом, блок модуляции с его восходящими и нисходящими связями работает по принципу обратной связи как единый саморегулирующий аппарат, который обеспечивает изменения тонуса коры и вместе с тем сам находится под его контролем. Этот аппарат используется для пластичного приспособления организма к условиям среды. Он содержит два источника активации: внутренний и внешний. Первый связан с обменными процессами, обеспечивающими гомеостаз, а второй – с воздействиями внешней среды. Первый, например задний гипоталамус, обеспечивает поведенческую активацию. Второй источник активации связан с воздействиями раздражителей внешней среды. Часть непрерывного потока сенсорных сигналов, поставляемых в кору специфическими системами, по коллатералям поступает в РФ ствола. В аппарате восходящей РФ формируется механизм преобразования сенсорной информации в две формы активации: тоническую (генерализованную) и фазическую (локальную). Тоническая форма активации связана с функцией нижних стволовых отделов РФ. Она генерализованно, диффузно поддерживает определенный уровень возбудимости в коре и подкорковых образованиях. Фазическая форма активности связана с верхними отделами ствола мозга и прежде всего с неспецифической системой таламуса. Неспецифическая система таламуса локально и избирательно распределяет воздействия восходящей активации на подкорковые образования, старую и новую кору.
Кроме того, установлено, что кора головного мозга наряду со специфическим функциональным вкладом оказывает «неспецифические» активирующие и тормозные влияния на нижележащие нервные образования. Корковые влияния, поступающие по нисходящим волокнам, представляют достаточно дифференцированную организацию и могут рассматриваться в качестве третьего источника активации. Наиболее обширные активирующие и инактивирующие избирательные влияния на ствол мозга исходят из лобных отделов коры.
3) Блок программирования, запуска и контроля поведенческих актов представляет собой сложноорганизованную, многоуровневую, построенную по принципу иерархии систему различных образований ЦНС.
Замысел движениярождается в ассоциативных лобных отделах новой коры; при участии моторной коры, базальных ганглиев, таламуса и мозжечка создается программа движения. Создание программы движения осуществляется по принципу доминанты. При этом происходит отбор структур, которые будут необходимы для организации движения в конкретном поведенческом акте. Это делается путем формирования соответствующего тонуса двигательных ядер ствола и спинного мозга, одновременно устанавливается необходимое соответствие с тонусом двигательных ядер тонуса нервных центров, ответственных за вегетативное обеспечение поведения. Собственно движениеосуществляется с участием двигательных единиц (ДЕ). Двигательная единица – мотонейрон спинного мозга и иннервируемая им группа мышечных волокон, образующих мышцу. Данный блок и все его узлы осуществляют свою деятельность на основе приема различного рода афферентаций (от зрительной сенсорной системы, вестибулярной, слуховой, тактильной, проприоцептивной, включая информацию от рецепторов растяжения мышц и т.д.).
Дата добавления: 2016-07-27; просмотров: 3588;
Поиск по сайту
Узнать еще
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- I.2. Антигены системы АВ0. Генетика. Структура
- I.2.1 ПЕРВЫЙ ЗАКОН НЬЮТОНА. ИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА.
- I.2.1 ПОЛНАЯ И ВНУТРЕННЯЯ ЭНЕРГИЯ СИСТЕМЫ. ТЕПЛОТА И РАБОТА
- I.3. Антитела системы АВ0
- I.5.4 НЕИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА. СИЛЫ ИНЕРЦИИ
- II. ЭЛЕКТРИЧЕСКИЙ ДИПОЛЬ. ДИПОЛЬНЫЙ МОМЕНТ СИСТЕМЫ ЭЛЕКТРИЧЕСКИХ ЗАРЯДОВ
- III. Избирательные системы.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
