Функциональная гистология коры больших полушарий
Кора головного мозга является высшим отделом центральной нервной системы. Она представляет собой тонкий слой нервной ткани, образующий множество складок. Общая площадь поверхности коры 2200 см2. Толщина коры колеблется от 1,3 до 4,5 мм. Объем коры равен примерно 600 см3. В состав коры головного мозга входит 109 – 1010 нейронов и еще большее число глиальных клеток (рис. 2.9). В пределах коры наблюдается чередование слоев, содержащих преимущественно тела нервных клеток, со слоями, образованными в основном их аксонами, и поэтому на свежем срезе кора головного мозга выглядит полосатой. На основании формы и расположения нервных клеток в коре с типичным строением можно выделить шесть слоев; некоторые из них подразделяются на два или более вторичных слоев. На основании строения коры выделяют следующие основные зоны: новую кору (неокортекс), старую кору (архикортекс), древнюю кору (палеокортекс) и межуточную кору (периархикортикальную и перипалеокортикальную). Самая обширная зона коры – неокортекс. Неокортекс занимает дорсальную и латеральную поверхность больших полушарий, в то время как палеокортекс находится на базальной и медиальной поверхности полушарий.
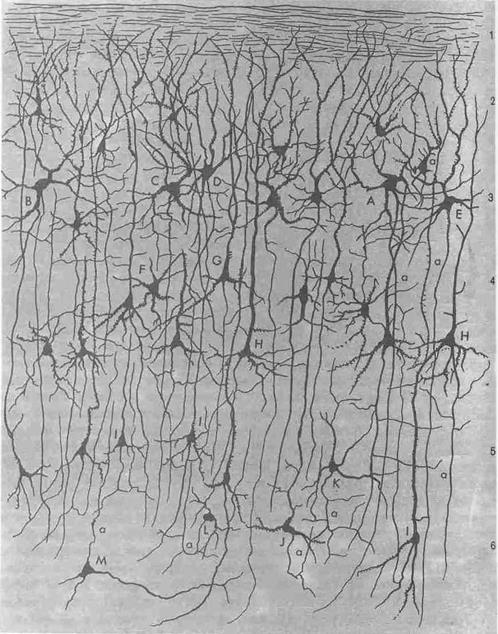
Рис. 2.9. Клеточный состав и слои коры головного мозга
В неокортексе различают следующие слои:
I. Молекулярный слой (плексиформный). В этом слое имеется множество волокон, образующих густое тангенциальное поверхностное сплетение, однако в нем мало клеток. Здесь содержатся в основном звездообразные мелкие клетки, которые осуществляют местную интеграцию деятельности эфферентных нейронов.
II. Наружный зернистый слой. Содержит мелкие нейроны различной формы, которые имеют синаптические связи с нейронами молекулярного слоя на всем поперечнике коры. В его глубине располагаются малые пирамидные клетки.
III. Наружный пирамидный слой. Этот слой состоит из пирамидных клеток малой и средней величины. Некоторые отделы коры в этом слое содержат крупные пирамидные клетки. Особенно много крупных пирамидных клеток в коре передней центральной извилины. Часть отростков этих клеток достигает первого слоя, участвуя в формировании тангенциального подслоя, другие погружаются в белое вещество полушарий мозга, поэтому III слой иногда обозначают как третичный ассоциативный.
IV. Внутренний зернистый слой. Характеризуется рыхлым расположением мелких нейронов различной величины и формы с преобладанием звездчатых, имеющих дугообразные возвратные аксоны. Аксоны клеток проникают в выше- и нижележащие слои. Звездчатые клетки представляют систему переключений с афферентных на эфферентные нейроны III и IV слоев. В IV слое также образуется тангенциальная прослойка из нервных волокон. Поэтому иногда этот слой обозначается как вторичный проекционно-ассоциативный. Внутренний зернистый слой является местом окончания основной массы проекционных афферентных волокон.
V. Внутренний пирамидный слой, или слой узловых клеток. Состоит в основном из средних и больших пирамидных клеток. Эти нейроны обладают длинными апикальными дендритами, простирающимися вплоть до молекулярного слоя, а также базальными дендритами, распространяющимися более или менее тангенциально по отношению к поверхности. Эти слои четко выражены в передней центральной извилине и незначительно – в других участках коры. Из этого слоя в основном формируются двигательные произвольные пути (проекционные эфферентные волокна).
VI. Слой веретеновидных клеток (полиморфный, или мультиформный). В этом слое расположены преимущественно веретеновидные нейроны, которые имеют короткие извитые верхушечные дендриты, заканчивающиеся в V и IV слоях коры. Аксоны многих клеток слоя объединяются в возвратные волокна, проникая в V слой. Глубинная часть этого слоя переходит в белое вещество (рис. 2.10).
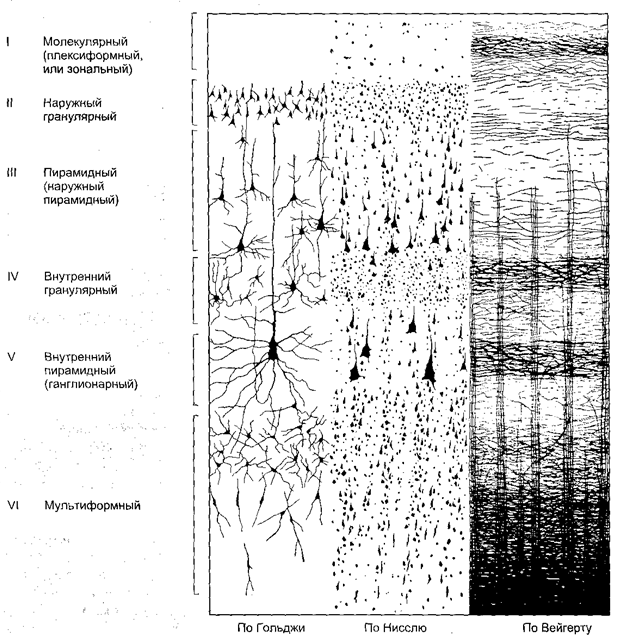
Рис. 2.10. Слоистое строение коры головного мозга
Нужно заметить, что нейроны каждого коркового поля имеют свои особенности строения. Цитоархитектонические слои состоят из нервных и глиальных клеток (олигодендро-, астромакроглии) и многочисленных нервных волокон. Нервные волокна образуют плотные сплетения, называемые нейропилем. Нервные клетки по своей форме весьма разнообразны. Отмечают до 56 разновидностей клеток коры. Более обобщенно выделяют наиболее многочисленные пирамидные нейроны (гигантские Беца, крупные моторные, средние, мелкие), звездчатые и веретенообразные. Доля пирамидных клеток среди всех нейронов коры колеблется от 51 до 86%, звездчатых клеток – от 8 до 47%, веретенообразных нейронов – от 2 до 6% (рис. 2.9).
Функционально в коре выделяют преимущественно возбуждающие нейроны: пирамидные, звездчатые, клетки Мартинотти (перевернутые пирамиды), глиеподобные и преимущественно тормозные: большие корзинчатые, малые корзинчатые, вертикально ориентированные, веретенообразные. Соединения между нейронами обеспечивают многочисленные синапсы и электротонические контакты. Большое значение в деятельности коры имеют шипиковые синапсы. Так, при развитии животных в условиях обогащенной раздражителями среды, по сравнению с сенсорной депривацией, происходит увеличение количества шипиков на дендритах. Умственная отсталость, снижение способности к обучению при хромосомных аберрациях у человека сопровождается уменьшением количества шипиков. Электротонические контакты осуществляются в коре в 20% случаев. Кроме того, в коре описаны несинаптические контакты между нейронами, функциональное назначение такого рода контактов остается еще неясным. В слоях I, II преимущественно имеются дендро-шипиковые контакты, в слоях III, IV – дендро-дендритические и сомато-дендритические, в слое V – сомато-сома-
тические и дендро-дендритические.
Американским физиологом В. Маунткаслом был выдвинут модульный принцип организации нейронов коры. Этот принцип базируется на трех отправных точках.
1. Кора головного мозга состоит из сложных многочисленных ансамблей, основная единица которых образована примерно сотней вертикально связанных нейронов всех слоев коры. Данный ансамбль называется мини-колонкой. В эти мини-колонки входят: а) нейроны, которые получают входные нейроны в основном от подкорковых структур, – например, от специфических сенсорных и моторных ядер таламуса; б) нейроны, получающие входные сигналы от других областей коры; в) все нейроны локальных сетей, образующих вертикальные клеточные колонки; г) клетки, передающие выходные сигналы от колонки назад к таламусу, другим областям коры, а иногда и клеткам лимбической системы.
2. Несколько таких сходных в своей основе простых вертикальных ансамблей могут объединяться с помощью межколоночных связей в более крупную единицу, перерабатывающую информацию, – модуль, или модульную колонку. Несмотря на различную плотность нейронов в слоях разных частей коры, общая структура и функции таких модульных колонок однотипны. Эти колонки различаются лишь по источнику получаемых ими входных сигналов и по мишеням, которым адресуются их выходные сигналы.
3. Маунткасл считает, что модули не только получают и перерабатывают информацию, но и совместно функционируют в составе обширных петель, по которым информация, выходя из колонок, передается другим кортикальным и субкортикальным мишеням, а затем возвращается обратно в кору. Эти петли обеспечивают упорядоченное поступление информации в кортикальные ансамбли.
Связи неокортекса
В новой коре выделяют несколько типов эфферентных и афферентных связей.
Эфферентными волокнами (кортико-фугальными) могут быть:
1) проекционные волокна к подкорковым образованиям (пути: кортико-спинальные, кортико-таламические, кортико-понтийные);
2) ассоциативные волокна, которые идут к одноименным и соседним областям коры одноименного полушария;
3) комиссуральные волокна, которые соединяют области коры обоих полушарий. Основными комиссурами являются мозолистое тело (corpus callosum) и передняя таламическая спайка. Мозолистое тело содержит очень много волокон. Например, у кошек на 1 мм2 приходится около 700 тысяч волокон.
Афферентными волокнами (кортико-петальными) являются ассоциативные, комиссуральные и таламокортикальный путь – главный афферентный путь к коре от подкорковых образований.
Афферентные волокна заканчиваются в основном в I-IV слоях коры. Исходя из этого, можно предположить, что в процессе обработки информации поверхностные слои ответственны главным образом за восприятие и обработку кортико-петальных сигналов. Особенное значение в этом процессе принадлежит IV слою коры.
Тела важнейших эфферентных нейронов коры лежат преимущественно в более глубоких слоях V-VI. Их считают зоной начала эфферентных путей коры.
Дата добавления: 2016-07-27; просмотров: 6982;
Поиск по сайту
Узнать еще
- А Нейропсихологические синдромы поражения задних отделов коры больших полушарий головного мозга.
- Акцентирование Больших Пальцев
- Аналитико-синтетическая деятельность коры больших полушарий
- АСИММЕТРИЯ ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА ЧЕЛОВЕКА
- Базальные ганглии. Морфофункциональная организация. Функции
- БАЗАЛЬНЫЕ ЯДРА КОРЫ БОЛЬШИХ ПОЛУШАРИЙ
- Базальные ядра. БЕЛОЕ ВЕЩЕСТВО полушарий головного мозга. ВНУТРЕННЯЯ КАПСУЛА. Боковые желудочки. ОБОНЯТЕЛЬНЫЙ МОЗГ. ЛИМБИЧЕСКАЯ СИСТЕМА
- Белки – структурно-функциональная основа жизни

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
