КАТАБОЛИЗМ ЖИРНЫХ КИСЛОТ
В живых организмах катаболизм ЖК протекает как в ферментативных так и в неферментативных реакциях.
· Ферментативный катаболизм ЖК происходит в основном в реакциях β-окисления. К побочным путям относиться ферментативное α- и ω-окисление ЖК, а также деградация ЖК в пероксисомах. Хотя эти побочные пути количественно менее важны, их нарушение может приводить к тяжелым заболеваниям.
· Неферментативный катаболизм ЖК протекает в реакциях перекисного окисления липидов (ПОЛ).
β-окисление ЖК
β-окисление — специфический путь катаболизма ЖК с неразветвленной средней и короткой углеводородной цепью. β-окисление протекает в матриксе митохондрий, при котором от С конца ЖК последовательно отделяется по 2 атома С в виде Ацетил-КоА. β-окисление ЖК происходит только в аэробных условиях и является источником большого количества энергии.
β-окисление ЖК активно протекает в красных скелетных мышцах, сердечной мышце, почках и печени. ЖК не служат источником энергии для нервных тканей, так как ЖК не проходят через гематоэнцефалический барьер, как и другие гидрофобные вещества.
β-окисление ЖК увеличивается в постабсорбтивный период, при голодании и физической работе. При этом концентрация ЖК в крови увеличивается в результате мобилизации ЖК из жировых ткани.
Активация ЖК
Активация ЖК происходит в результате образования макроэргической связи между ЖК и HSКоА с образованием Ацил-КоА. Реакцию катализирует фермент Ацил-КоА синтетаза:
RCOOH + HSKoA + АТФ → RCO~SКоА + АМФ+ PPн
Пирофосфат гидролизуется ферментом пирофосфатазой: Н4Р2О7 + Н2О → 2Н3РО4
Ацил-КоА синтетазы находятся как в цитозоле (на внешней мембране митохондрий), так и в матриксе митохондрий. Эти ферменты отличаются по специфичности к ЖК с различной длиной углеводородной цепи.
Транспорт ЖК
Транспорт ЖК в матрикс митохондрий зависит от длины углеродной цепи.
ЖК с короткой и средней длиной цепи (от 4 до 12 атомов С) могут проникать в матрикс митохондрий путём диффузии. Активация этих ЖК происходит ацил-КоА синтетазами в матриксе митохондрий.
ЖК с длинной цепью, сначала активируются в цитозоле (ацил-КоА синтетазами на внешней мембране митохондрий), а затем переносятся в матрикс митохондрий специальной транспортной системой с помощью карнитина. Карнитин поступает с пищей или синтезируется из лизина и метионина с участием витамина С.
· В наружной мембране митохондрий фермент карнитинацилтрансфераза I (карнитин-пальмитоилтрансфераза I) катализирует перенос ацила с КоА на карнитин с образованием ацилкарнитина;
· Ацилкарнитин проходит через межмембранное пространство к наружной стороне внутренней мембраны и транспортируется с помощью карнитинацилкарнитинтранслоказы на внутреннюю поверхность внутренней мембраны митохондрий;
· Фермент карнитинацилтрансфераза II катализирует перенос ацила с карнитина на внутримитохондриальный HSКоА с образованием Ацил-КоА;
· Свободный карнитин возвращается на цитозольную сторону внутренней мембраны митохондрий той же транслоказой.
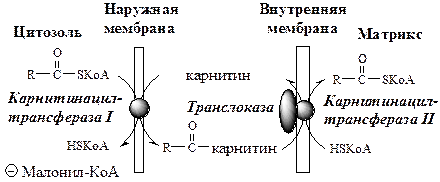
Реакции β-окисление ЖК
1. β-окисление начинается с дегидрирования ацил-КоА ФАД-зависимой Ацил-КоА дегидрогеназой с образованием двойной связи (транс) между α- и β-атомами С в Еноил-КоА. Восстановленный ФАДН2 окисляясь в ЦПЭ, обеспечивает синтез 2 молекул АТФ;
2. Еноил-КоА гидратаза присоединяет воду к двойной связи Еноил-КоА с образованием β-оксиацил-КоА;
3. β-оксиацил-КоА окисляется НАД зависимой дегидрогеназой до β-кетоацил-КоА. Восстановленный НАДН2, окисляясь в ЦПЭ, обеспечивает синтез 3 молекул АТФ;
4. Тиолаза с участием HКоА отщепляет от β-кетоацил-КоА Ацетил-КоА. В результате 4 реакций образуется Ацил-КоА, который короче предыдущего Ацил-КоА на 2 углерода. Образованный Ацетил-КоА окисляясь в ЦТК, обеспечивает синтез в ЦПЭ 12 молекул АТФ.
Затем Ацил-КоА снова вступает в реакции β-окисления. Циклы продолжаются до тех пор, пока Ацил-КоА не превратится в Ацетил-КоА с 2 атома С (если ЖК имела четное количество атомов С) или Бутирил-КоА с 3 атомами С (если ЖК имела нечетное количество атомов С).
Дата добавления: 2016-07-27; просмотров: 3232;
Поиск по сайту
Узнать еще
- А) кислотные свойства
- А. ОДНООСНОВНЫЕ ПРЕДЕЛЬНЫЕ КИСЛОТЫ
- Аденилат (адениловая кислота), Гуанилат (гуаниловая кислота),
- АДЕНОЗИНТРИФОСФОРНАЯ КИСЛОТА
- Азотная кислота и ее свойства.
- Активация аминокислот при биосинтезе белка.
- АМИДЫ СУЛЬФАНИЛОВОЙ КИСЛОТЫ (СУЛЬФАНИЛАМИДНЫЕ ПРЕПАРАТЫ)
- АМИНОКИСЛОТНЫЕ СМЕСИ И БЕЛКОВЫЕ ГИДРОЛИЗАТЫ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
