Механизм мышечного сокращения.
В соответствии с теорией скользящих нитей мышца сокращается в результате укорочения множества последовательно соединенных саркомеров в миофибриллах, при этом тонкие актиновые нити скользят вдоль толстых миозиновых, двигаясь между ними к середине саркомера.
Миозиновые нити несут поперечные выступы с головками, состоящими примерно из 150 молекул миозина (рисунок 7.6А). Во время сокращения каждая головка (поперечный мостик) может связывать миозиновую нить с соседними актиновыми. Движение головок создаёт объединённое усилие, как бы «гребок», продвигающий актиновые нити в середину саркомера [31].
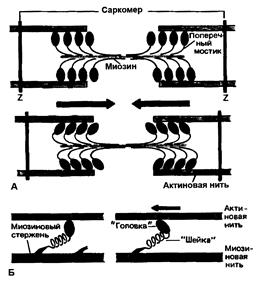
Рисунок 7.6 Функция поперечных мостиков.
А – модель механизма сокращения: миозиновая нить с поперечными мостиками, прикреплёнными к соседним актиновым нитям. Вверху – до, внизу – после «гребкового» движения.
Б – модель механизма генерирования силы при статическом напряжении. Слева - до, справа – после «гребка» (по Й. Рюэгг[31]).
В расслабленном состоянии механизм взаимодействия миозиновых головок с актиновыми нитями блокируется за счёт того, что участок актиновых нитей, предназначенный для связывания с головками миозина, перекрыт молекулами белков тропонина и тропомиозина.
Мышечное сокращение происходит под воздействием двигательного нервного импульса, представляющего собой волну повышенной мембранной проницаемости, распространяющейся по нервному волокну. Эта волна передаётся на Т-систему саркоплазматической сети и в конечном итоге достигает цистерн, содержащих ионы кальция. Проницаемость стенок цистерн повышается, ионы кальция выходят из цистерн в саркоплазму, где их концентрация за очень короткое время (около 3милисекунд) увеличивается примерно в 1000 раз. Ионы кальция, взаимодействуя с тропонином, изменяют его пространственную форму, что вызывает смещение молекул тропомиозина, обеспечивая доступ миозиновых головок к активным участкам актиновых нитей. В результате этого между миозином и актином возникает поперечный мостик, расположенный под углом 90о (рисунок 7.6А). Поскольку в толстые и тонкие нити входит большое число молекул миозина и актина, между мышечными нитями образуется довольно большое число поперечных мостиков. Образование связи между актином и миозином сопровождается повышением АТФ-азной активности миозина, разрешая её расщепление. За счёт энергии, выделяющейся при гидролизе АТФ, миозиновая головка подобно веслу лодки поворачивается и мостик между толстыми и тонкими нитями оказывается под углом 45о, что приводит к скольжению мышечных нитей навстречу друг другу (рисунок 7.6А) [11].
Совершив поворот, мостики между тонкими и толстыми нитями разрываются, АТФ-азная активность миозина резко снижается и гидролиз АТФ прекращается. Но если двигательный нервный импульс продолжает поступать в мышцу и в саркоплазме сохраняется высокая концентрация ионов кальция, поперечные мостики образуются снова, повторно включается гидролиз АТФ, дающий энергию для поворота поперечных мостиков с последующим их разрывом. За счёт ритмичных прикреплений и отделений миозиновых головок актиновая нить подтягивается к середине саркомера подобно тому, как группа людей тянет верёвку, перебирая её руками. Когда принцип «вытягивания верёвки» реализуется во множестве последовательно расположенных саркомеров, повторяющиеся молекулярные движения поперечных мостиков приводят к движению всей мышцы [31].
Каждый цикл сокращения, включающий образование, поворот и разрыв мостика, требует расходования одной молекулы АТФ в качестве источника энергии [11].
При сокращении мышцы происходит её укорочение. Но при статической нагрузке длина мышцы не изменяется. Что же в таком случае происходит в мышце с точки зрения «гребковой» теории Хаксли?
Благодаря упругости поперечных мостиков саркомер может развивать силу даже без скольжения нитей относительно друг друга. Процесс генерирования изометрической (т.е. статической) силы показан на рисунке 7.6Б. Сначала головка миозиновой молекулы прикрепляется к актиновой нити под прямым углом. Затем она наклоняется под углом 45о, возможно благодаря притяжению между соседними точками прикрепления на ней и на актиновой нити. При этом головка действует как миниатюрный рычаг, приводя внутреннюю упругую структуру поперечного мостика в напряжённое состояние. Упругое напряжение, создаваемое индивидуальным поперечным мостиком очень мало и для создания необходимой мышечной силы требуется объединение усилий огромного количества таких соединённых параллельно мостиков. Они будут тянуть соседние актиновые нити, как команда игроков тянет канат. Но даже при изометрическом сокращении поперечные мостики не находятся в непрерывно напряжённом состоянии. Каждая миозиновая головка уже через десятые или даже сотые доли секунды отделяется от актиновой нити, прикрепляясь к ней снова через такое же короткое время. Однако несмотря на чередование прикреплений и отделений, следующих с частотой от 5 до 50 раз в секунду, сила, развиваемая мышцей, остаётся неизменной, так как статистически в каждый момент времени в прикреплённом состоянии находится одно и то же количество мостиков [31].
Расслабление мышцы происходит после прекращения поступления двигательного импульса. При этом ионы кальция переходят обратно в цистерны саркоплазматического ретикулума. Уборка ионов кальция происходит в сторону с более высокой концентрацией этих ионов, поэтому этот процесс требует дополнительных затрат энергии. Процесс уборки ионов кальция называют кальциевым насосом и производится он за счёт энергии, получаемой при расщеплении АТФ, причём на уборку каждого иона кальция затрачивается две молекулы АТФ [24]. Снижение концентрации кальция в саркоплазме вызывает изменение пространственной ориентации тропонина, что в конечном итоге приводит к невозможности образования поперечных мостиков между толстыми и тонкими нитями. За счёт упругих сил, возникших ранее (при мышечном сокращении) в коллагеновых нитях, окружающих мышечное волокно, мышца расслабляется и возвращается в исходное положение, чему также может способствовать и сокращение мышц-антогонистов [11].
Дата добавления: 2020-10-01; просмотров: 608;
Поиск по сайту
Узнать еще
- A) четырехшарнирный механизм
- III. Механизм действия ионизирующих излучений на биологические структуры
- III. Механизмы, средства и методы психологического воздействия.
- R-плазмиды, функции, строение. Пути передачи. Механизм множественной лекарственной устойчивости.
- VI.НЕЙРОХИМИЧЕСКИЕ МЕХАНИЗМЫ ПЛАСТИЧНОСТИ И ПАМЯТИ.
- Алгоритм проектирования кулачкового механизма по допустимому углу давления
- Аллергические реакции 1 типа (реагиновые). Стадии, медиаторы аллергии 1 типа, механизмы их действия. Клинические проявления (анафилактический шок, атопические реакции).
- АНАЛИЗ МЕХАНИЗМА ЦЕНООБРАЗОВАНИЯ.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
