Строение и химический состав скелетных мышц
Скелетная мышца состоит из мышечных волокон (миоцитов). Мышечные волокна представляют собой гигантские многоядерные клетки длиной от 0,1 до 2-3 сантиметров, а в некоторых мышцах миоциты достигают 12 сантиметров. Площадь поперечного сечения мышечных клеток составляет от 3 до 10 квадратных микрометров.
Волокно покрыто эластичной оболочкой — сарколеммой и состоит из саркоплазмы, структурными элементами которой являются такие органоиды, как митохондрии, рибосомы, трубочки и пузырьки саркоплазматической сети (ретикулума) и так называемая Т-система а также различные включения. В саркоплазме условно выделяют две части – саркоплазматический матрикс и саркоплазматический ретикулум.
Саркоплазматический ретикулум, представляющий собой определённым образом организованную сеть соединяющихся цистерн (содержащих в большой концентрации ионы кальция) и трубочек, играет важную роль в механизмах сокращения и расслабления мышцы. Кроме того, к части ретикулума прикреплены рибосомы, специальные сферические образования, на которых и при участии которых происходит биосинтез белков. Саркоплазматическая сеть с помощью особых трубочек, называемых Т-системой, связана с оболочкой мышечной клетки. Т-система также имеет прямое отношение к мышечному сокращению, так как по ней передаётся изменение электрического потенциала поверхностной мембраны элементам ретикулума, что приводит к освобождению ионов кальция, поступающих к миофибриллам и запускающих процесс мышечного сокращения [24].
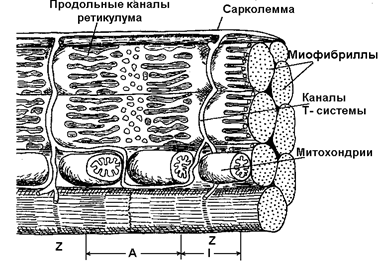
Рисунок 7.3 Строение Т-системы и саркоплазматического ретикулума мышечного волокна
[из [24] по: Кроленко, 1975].
Саркоплазматический матрикс представляет собой коллоидный раствор, содержащий белки, гликоген, жировые капли и другие включения [11]. Миофибриллы – сократительные элементы мышечных клеток – также находятся в саркоплазматическом матриксе.
Кроме того, в саркоплазме находятся ферменты гликолиза, расщепляющие гликоген или глюкозу до пировиноградной или молочной кислоты и креатинкиназа – фермент, ускоряющий креатинфосфатную реакцию. Особый белок саркоплазмы – миоглобин – обеспечивает некоторый запас кислорода в мышечной ткани, а также участвует в переносе кислорода от сарколеммы к митохондриям.
Мышечная клетка имеет не одно, а множество ядер, которые располагаются на её периферии – под сарколеммой. Внутри каждого ядра находится ДНК, являющаяся носителем носледственной информации и состоящая из генов, в которых закодирована структура всех синтезируемых мышечными волокнами белков.
Лизосомы, представляющие собой микроскопические пузырьки, содержат в растворённом виде различные ферменты, способные в условиях кислой реакции среды расщеплять различные высокомолекулярные вещества. Такая необходимость может возникать в мышечных клетках, например, при очень напряжённой мышечной деятельности.
Митохондрии
Митохондрии, одни из важнейших структурных компонентов мышечного волокна, располагаются цепочками вдоль миофибрилл (рисунок 7.3), тесно соприкасаясь с мембранами ретикулума. В митохондриях протекает аэробное окисление углеводов, жиров и аминокислот, а за счёт энергии, выделяющейся при окислении, происходит ресинтез АТФ.
Митохондрии ограничены двумя мембранами (рисунок 7.4). Наружняя митохондриальная мембрана имеет ровные контуры, не образует выпячиваний или складок. Наружную мембрану от внутренней отделяет межмембранное пространство. Внутренняя мембрана ограничивает внутреннее содержимое митохондрии, ее матрикс. Характерной чертой внутренней мембраны митохондрий является их способность образовывать многочисленные выпячивания внутрь митохондрий. Такие выпячивания чаще всего имеют вид плоских гребней, или крист, существенно увеличивая поверхность внутренней мембраны. Мембраны митохондрий построены из белка и содержащих фосфорную кислоту жироподобных веществ - фосфолипидов. На внутренней мембране в определённом порядке расположены биологические катализаторы – ферменты, при помощи которых происходят окислительные процессы, а также компоненты дыхательной цепи – главной системы превращения энергии в митохондриях. На внешней мембране митохондрий в определённом порядке расположены ферменты, не имеющие отношения к дыхательной цепи. Немало ферментов в растворённом виде содержится и в матриксе. Кроме того, матрикс митохондрий содержит рибосомы и митохондриальную ДНК.
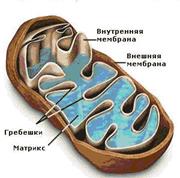
Рисунок 7.4Схема строения митохондрии
(по А.Кузнецов, [29])
Великое множество миофибрилл, содержащихся в мышечных волокнах, требуют большого количества АТФ, которое должно быть доставлено к каждому саркомеру миофибрилл. На продольных ультратонких срезах скелетных мышц в электронном микроскопе видны многочисленные округлые мелкие сечения митохондрий, располагающихся в соседстве с саркомерами. Если же исследовать поперечные срезы мышечных волокон на уровне Z-дисков (см. п.7.2.1.2), то видно, что мышечные митохондрии представляют собой не мелкие шарики или палочки, а как бы паукообразные структуры, отростки которых могут ветвиться и простираться на большие расстояния, иногда через весь поперечник мышечного волокна. При этом разветвления митохондрий окружают каждую миофибриллу в мышечном волокне, снабжая их АТФ, необходимой для мышечного сокращения. Следовательно, в плоскости z-диска митохондрии представлены типичным митохондриальным ретикулумом – единой митохондриальной системой. Такой пласт или этаж митохондриального ретикулума повторяется дважды на каждый саркомер, а все мышечное волокно имеет тысячи поперечно расположенных поэтажных пластов митохондриального ретикулума. Было обнаружено, что между этажами вдоль миофибрилл располагаются нитчатые митохондрии, соединяющие эти митохондриальные пласты. Тем самым создается трехмерная картина митохондриального ретикулума, проходящего через весь объем мышечного волокна [28].
Предполагается, что с помощью специальных межмитохондриальных соединений или контактов может происходить функциональное объединение отдельных митохондрий и митохондриальных ретикуломов в единую энергетическую систему, позволяющую всем миофибриллам в мышечном волокне сокращаться синхронно по всей длине, поскольку механизм взаимодействия митохондрий посредством межмитохондриальных контактов может обеспечить синхронное поступление АТФ во все участки сокращающегося мышечного волокна.
Механизм кооперации и синхронизации работы митохондрий позволяет вести синтез АТФ в любой точке поверхности внутренней мембраны таких разветвлённых митохондрии, обеспечивая энергией для сокращения те участки мышечного волокна, где в этом возникает необходимость. Но связывание отдельных митохондрий в единую цепь с помощью межмитохондриальных контактов наряду с очевидными преимуществами имеет и существенный недостаток. Дело в том, что при функциональном объединении митохондрий в единую митохондриальную систему любое существенное повреждение (пробой) её внутренней мембраны приводит к потере способности к ресинтезу АТФ сразу у всей объединённой группы митохондрий.
При проведении серии развивающих тренировок по подтягиванию направленных на развитие статической выносливости мышц-сгибателей кисти нередко используется метод выполнения нагрузки «до отказа». Если тренировки разделены недостаточным для восстановления интервалом отдыха, после проведения 4-5 развивающих тренировок подряд, в ходе которых может наблюдаться существенный прирост времени виса (т.е. увеличение аэробных возможностей мышц), неожиданно наступает срыв адаптации и возврат времени виса к первоначальному уровню.
Например, если спортсмен форсирует тренировочный процесс и выполняет через день по 4-6 подходов до отказа, подняв за 2 недели вис с подтягиванием в темпе 1 раз в 8 секунд с 2 до 4 минут (такое возможно у квалифицированных спортсменов, например, после длительного вынужденного перерыва в тренировках), то внезапно – без видимых причин - время виса может упасть до прежних двух минут и даже меньше.
Долгое время было непонятно, почему так происходит. В качестве одной из возможных причин называлась перегрузка нервной системы тренировками до отказа. Но срыв адаптации обычно происходил на фоне эмоционального подъёма от быстрого прогресса тренировочных результатов и связанного с этим желания тренироваться всё больше и больше и имел мало общего с нервным срывом.
Возможно, что резкое падение результатов происходит из-за пробоя внутренней мембраны митохондриальной системы мышечного волокна, вследствие, например, чрезмерного закисления мышц на предшествующей срыву адаптации тренировке. В этом случае повреждение небольшого по площади участка любой из митохондрий, входящих в митохондриальную сеть, должно приводить к отключению механизма аэробного окисления сразу во всей сети.
Тренировки с облегчением в 5-7% от веса тела позволяют резко (в 1,5 – 2 раза) увеличить объём тренировочной работы за счёт увеличения количества подтягиваний в подходе с соответствующим увеличением времени выполнения подхода. При этом энергопродукция смещается в сторону аэробного окисления, всё в большей степени активизируя работу митохондриальной системы. Серия развивающих тренировок с облегчением без должного интервала отдыха между ними также может привести к скачкообразному падению результатов, что также может быть объяснено повреждением внутренних мембран митохондрий продуктами метаболизма.
Можно ли каким-либо образом почувствовать приближение момента срыва адаптации и, снизив нагрузку, предотвратить это нежелательное явление?
Биологическое окисление, протекающее в митохондриях, состоит в окислении органических субстратов, например глюкозы, до углекислого газа и воды с выделением около 680 ккал (в расчёте на 1 моль, т.е. 180 г глюкозы), которая в дальнейшем идёт на создание макроэнергетической связи в молекуле АТФ (фосфорилирование АДФ). Окисление и фосфорилирование – это два, в принципе, независимых процесса, которые для эффективного ресинтеза АТФ должны быть сопряжены. Сопряжение окисления и фосфорилирования происходит на внутренних мембранах митохондрий. Поэтому, когда мембраны повреждены, происходит разобщение этих процессов. Реакции окисления глюкозы продолжают идти, а ресинтез АТФ замедляется или прекращается. И сейчас даже неважно, что является причиной повреждения мембран – избыток молочной кислоты, недостаток кислорода или повышенное его потребление, свободнорадикальное окисление или это происходит по каким-то иным причинам. Важно, что при повреждении мембран митохондрий в результате чрезмерного воздействия тренировочных нагрузок нарушается процесс ресинтеза АТФ, а энергия, выделяющаяся в процессе биологического окисления, теперь может превращаться только в тепло, приводя к локальному нагреву мышечной ткани.
Но одним из отдалённых признаком перетренированности является внезапная испарина, выступающая не только на лбу, но и на рабочих мышцах спортсмена в начале выполнения даже не очень напряжённой нагрузки. Возможно, таким образом организм реагирует на начинающийся процесс разобщения окисления и фосфорилирования, связанный с повреждением мембран митохондрий. Поэтому, если после серии развивающих нагрузок на очередной тренировке вы начинаете по непонятным причинам сильно потеть, стоит задуматься: а не перебрали ли вы с нагрузкой?
В любом случае безопасным (но в то же время обеспечивающим прогресс) считается вариант, когда тяжёлые развивающие тренировки проводятся не чаще одного раза в 5 – 7 дней.
Экспериментальные данные говорят в пользу того, что увеличение числа митохондрий происходит путём роста и деления предшествующих митохондрий. Более того, митохондрии обладают собственным генетическим аппаратом, т.е. обладают полной системой авторепродукции, хотя и находящейся под генетическим контролем со стороны клеточного ядра [28].
Все митохондрии в теле человека наследуются от матери, а не от отца, поэтому способность к длительному выполнению упражнений передаётся по материнской линии.
По форме и размеру митохондрии напоминают бактерий; они содержат собственную ДНК и размножаются делением. Эти и другие факты привели к возникновению гипотезы о том, что много миллионов лет назад бактерии проникли в более высокоразвитые клетки и прочно обосновались в них, потеряв былую самостоятельность и со временем превратившись в клеточные органеллы, которые теперь называют митохондриями [30].
Миофибриллы
Сократительные элементы – миофибриллы – занимают большую часть объёма мышечных клеток. Миофибриллы состоят из многочисленных параллельно расположенных нитей – филаментов. Перегородки, называемые Z – пластинками, разделяют их на отдельные участки, называемые саркомерами. Строение саркомера мышечного волокна показано на рисунке 7.5.
Мышечные нити – филаменты бывают двух типов: толстые и тонкие.Толстые имеют диаметр около 10 нм (1 нм = 10-9 м), а тонкие – около 5 нм. Толстые нити, состоящие из белка миозина, расположены в дисках А (рис. 7.3, 7.5), а тонкие, основным белком которых является актин, находятся в дисках I, частично заходя в диск А. Середину диска I пересекает Z – пластинка, соединяющая тонкие нити между собой и с сарколеммой. В поперечном сечении толстые и тонкие нити располагаются правильными шестиугольниками так, что каждая толстая нить окружена шестью тонкими, а каждая тонкая нить может вступать в контакт с тремя толстыми [24].
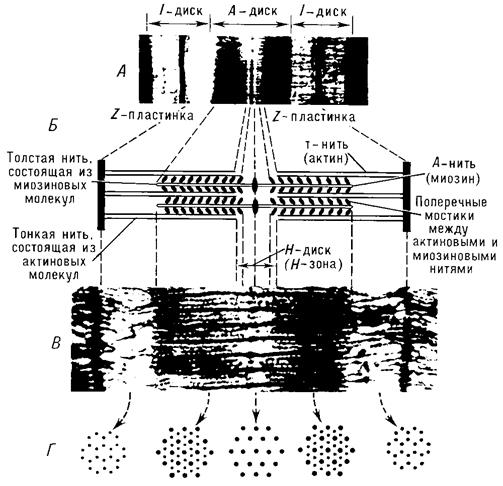
Рисунок 7.5. Строение саркомера поперечнополосатого мышечного волокна: А — электронная микрофотография (малое увеличение), на которой четко видна структура саркомера; Б — схема саркомера; В — электронная микрофотография с высокой разрешающей способностью; Г — поперечное сечение саркомера на различных уровнях, видно положение толстых и тонких нитей в различных участках покоящегося саркомера (по Х. Хаксли)
Из-за особенностей своих оптических свойств миозиновые нити, находящиеся в середине саркомера, выглядят в световом микроскопе тёмной полосой, а актиновые нити – светлой. Именно в результате такого периодического чередования светлых и тёмных полос в бесчисленных саркомерах миофибриллы выглядят поперечно-полосатыми.
Дата добавления: 2020-10-01; просмотров: 715;
Поиск по сайту
Узнать еще
- E) Расчет структурных составляющих очага деформации с одним нейтральным сечением
- F. Построение диаграмм сумм рангов
- II. СТРОЕНИЕ ОБЩЕСТВА
- II. Химический состав природных воды
- II.2. Химический (молекулярный) состав нефтей и природных газов
- III этап. Составление программного кода
- III.I. Слой, слоистость и строение слоистых толщ
- III.XI. Строение поверхностей несогласия

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
