Классификация экосистем и их изменчивость во времени
Общеизвестно положение о динамическом равновесии природных экосистем и условий окружающей среды. Однако механизм этого равновесия пока изучен недостаточно. Чаще всего рассуждения на эту тему идут на уровне соответствия растительности и почв климатическим условиям конкретной территории: в тропическом климате тропические экосистемы, в степном - степные, в бореальном - лесные. Когда же проблема парникового эффекта поставила остро вопрос о том, как будут реагировать земные экосистемы на потепление климата планеты, обнаружились серьезные пробелы знаний и противоречия суждений авторитетных специалистов. Механизм реагирования экосистем на изменения факторов среды в целом и в деталях оказался практически не изученным. Актуальность проблемы заставляет иначе смотреть на имеющийся запас знаний.
Главная новость для экологов в этом отношении заключается в том, что все природные экосистемы развиваются в режиме перманентной адаптации к постоянным изменениям факторов среды в суточных, сезонных, годовых, многолетних циклах. Каждая природная экосистема имеет свой диапазон оптимальных флуктуации, заложенный в генетическом аппарате отдельных особей. Отклонение реального хода условий от идеального, генетически обусловленного, сказывается на конечном итоге развития системы. Аномалии проявляются в изменении габитуса, массы, продуктивности экосистемы.
Даже в относительно стабильных условиях среды экосистема должна приспосабливаться к колебаниям этих условий в суточном, годовом и многолетнем циклах. Монотонный режим функционирования экосистемы не более, чем абстракция. Экосистема всегда стремится к равновесию с условиями среды, но никогда его не достигает потому, что условия среды меняются раньше и быстрее,
чем экосистема реагирует на эти изменения. Нормальным режимом функционирования любой экосистемы следует считать режим непрерывной (перманентной) адаптации к непрерывно меняющимся условиям среды. Реакция экосистемы зависит от величины и продолжительности изменений факторов среды.
Незначительные колебания факторов среды сопровождаются флуктуациями экосистемы - изменениями в пределах диагностических параметров таксона. Изменение массы экосистемы и ее компонентов, соотношения видов в биоценозе остаются в пределах, которые характеризуют данную экосистему.
Выход условий среды за пределы оптимального диапазона может изменить видовой состав биоценоза, его продуктивность, соотношение фракций экомассы до такой степени, что экосистема утратит часть прежних диагностических признаков и приобретет признаки другого таксона классификации. Виды, для которых новые условия неприемлемы, уступят место вида, для которых новые условия являются оптимальными. Такие изменения, в результате которых меняется таксономическое положение экосистемы, называются метаморфозами.
Маятниковый характер флуктуаций факторов среды приводит к тому, что каждая экосистема вынуждена попеременно реагировать то на улучшение, то на ухудшение условий среды, что компенсирует до некоторой степени результаты изменений. Однако реакция экосистемы на изменения факторов среды в сторону минимума и максимума различна. При улучшении условий адаптивная реакция системы не встречает сопротивления и эффект получается максимальный. Зато при ухудшении условий система включает буферные механизмы, которые сдерживают ее реакцию на неблагоприятную смену условий среды. После прохождения полного цикла флуктуации условий среды всегда получается положительный эффект, который выражается в увеличении биомассы, урожая плодов и семян.
Функционирование информационно-управляемых систем должно быть подчинено определенной цели, для достижения которой они имеют набор функциональных блоков, уровней управления и соответствующих уровней информации, корректирующих процесс достижения цели (управления). Целесообразность технических систем определяется человеком, поскольку всякая машина предназначена для достижения конкретной цели.
У природных систем цель - самовоспроизводство, которое заложено в генотипе. Ради воспроизводства себе подобных (репродукции) живые системы адаптируются в пределах генотипа к меняющимся условиям среды. Они могут изменить стереотип поведения, форму и окраску в зависимости от характера и степени от-
клонения реальных условий среды от оптимальных, обусловленных генетическим кодом. Важно отметить принципиальное различие реакции организма и сообщества на отклонение факторов среды от оптимального диапазона. Организм в ответ на смену условий изменяет режим функционирования, сохраняя структуру. Биоценоз изменяет структуру, сохраняя режим функционирования: одни виды сменяются другими более устойчивыми к новым условиям, а емкость и скорость круговорота вещества остаются прежними.
Примером согласованности функций компонентов экосистемы может служить ее реакция на флуктуации условий среды. Согласованность функций фитоценоза и педоценоза обусловлена адекватной реакцией автотрофных и гетеротрофных организмов на изменения гидротермических условий. Нарушают эту синхронность автономная реакция фитоценоза на свет, а педоценоза на аэрацию и значительная инерционность реакции педоценоза по сравнению с фитоценозом.
В идеальном случает педоценоз в каждый данный момент времени выделяет фитоценозу столько и таких минеральных элементов, сколько и каких ему требуется в данный момент для фотосинтеза. В реальных экосистемах такое случается редко вследствие указанных выше разногласий. Поэтому более характерны две крайние ситуации: педоценоз выделяет либо меньше минеральных элементов, чем требуется фитоценозу, либо больше. Количественное значение дефицита и избытка не так важно.
Если в результате флуктуаций факторов среды в почве образуется избыток минеральных элементов, включается механизм гумификации, вторичного синтеза органического вещества при взаимодействии минеральных ионов со свободными органическими радикалами. В случае дефицита минеральных элементов, фитоценоз сам провоцирует активизацию почвенной биоты. Он выделяет в ризосферу корневые выделения, которые провоцируют вспышку активности микрофлоры, минерализующей гумус прикорневой зоны, и ликвидируют дефицит минеральных элементов полностью или частично. Динамическое равновесие экосистемы заключается в периодической смене процессов накопления и высвобождения минеральных элементов в педоценозе и поддержания баланса элементов в экосистеме.
Согласно алгоритму развития экосистемы в режиме непрерывной (перманентной) адаптации к флуктуациям среды, количество минеральных элементов в процессе эволюции экосистем постепенно и неуклонно возрастает. Природа создала эффективный механизм защиты массы минеральных элементов от катает-
рофических потерь. Это ярко проявляется на примере пироген-ной сукцессии лесных экосистем.
Во время лесного пожара высвобождается большое количество зольных элементов, которые могут быть вынесены за пределы экосистемы водными и воздушными потоками. Спасают положение "сорные растения", которые при отсутствии конкурентов и обилии пищи способны увеличить свои габариты до гигантских размеров и поглотить максимум свободных минеральных элементов, связать их, защитить от выноса из экосисиемы. После отмирания сорняков минеральные элементы закрепляются в гумусе и остаются в запасном фонде экосистемы.
Все известные технологии земледелия базируются на приемах искусственного провоцирования минерализации почвенного гумуса с целью выделения дополнительных порций элементов минерального питания для культурных растений. Однако, в отличие от природных фитоценозов, наши монокультурные посевы потребляют только 20-30% выделенных с помощью плугов и других рыхлителей минеральных элементов. Остальная масса, как после пожара, обречена на вынос из экосистемы водными и воздушными потоками. Сорняки пытаются выполнить свою экологическую миссию, определенную природой, защитить минеральные элементы от нежелательных потерь, но земледельцы не позволяют им этого делать, уничтожают всеми способами и методами. С этой точки зрения беспахотное земледелие или система нулевой обработки почвы становится более экологичной, чем паро-пропашная, поскольку уменьшают количество лишних элементов питания.
Полидоминантные посевы значительно повышают коэффициент использования искусственно минерализованных элементов питания. Сочетание этих принципов с разработкой соответствующих технических средств и учетом экологического потенциала каждого конкретного угодья позволит снизить негативный эффект сельскохозяйственного воздействия на функции экосистем и позволит оздоровить среду обитания человека как биологического вида, преодолевшего естественный лимит численности популяции. В результате длительного и многоэтапного периода сукцессии, она вступает в очередную стадию климакса. В цикле развития экосистемы (экогенезе) стадия климакса - заключительная и наиболее продолжительная. Именно в этот период экосистема функционирует в полном объеме в режиме наиболее устойчивого динамического равновесия с факторами внешней среды. Сукцессии периодически прерывают эту стадию естественным образом исключительно для обновления экотопа. Поскольку растения не могут, как животные, кочевать по территории в поисках пищи, они вынуждены кочевать во времени. Мозаика
фитоценоза только кажется неизменной. На самом деле мозаичная структура фитоценозов сохраняет только соотношение разнокачественных пятен в растительном и почвенном покрове, однако сами пятна периодически меняются местами. Впечатление стабильности структуры фитоценоза создается вследствие различия характерных времен жизненного цикла экосистемы и наблюдателя. Об этом кратко говорилось в главе 1.2. Здесь мы остановимся более подробно на описании механизма циклических метаморфозов почвенно-растительного покрова в двух резко различных регионах: Западной Сибири и Прикаспийской низменности.
Западная Сибирь. Исследователей подзолистых и серых лесных почв с давних пор интересовало происхождение второго гумусового горизонта (A2h, Bh), встречающегося в некоторых почвенных профилях в виде темной полосы или серии темных пятен на контакте подзолистого и иллювиального горизонтов (Гордя-гин, 1901; Драницын, 1914; Глинка, 1923;,Горшенин и Сельская, 1929; Ильин, 1929,1937; Петров, 1937; Иванова и Двинских, 1944; Кузнецов, 1948 и многие другие). Особенно возрос этот интерес в 60-е годы в связи с появлением новых данных, характеризующих физико-химические свойства этого горизонта в почвах различных регионов, состав гумуса и его абсолютный возраст (Буди-на и Ерохина, 1969; Гаджиев, 1964; Герасимов, 1962; Добровольский и др., 1970; Ковалев и Гаджиев, 1968; Кузьмин, 1969; Пономарева, 1956; Пономарева и Толчельников, 1968; Толчельников, 1970; Уфимцева, 1965, 1970; Хантулев и др., 1969; Хисмату-лин,1970 и др.). Новая волна интереса к этому вопросу прошла в 80-е годы (Нечаева, 1985; Караваева и др., 1986).
Упомянутые авторы согласованно показывают, что A2h выделяется в почвенном профиле не столько количеством, сколько качеством гумуса, его гуматным составом. В гумусе A2h действительно преобладают гуминовые кислоты фракции 2, характеризующиеся высокой оптической плотностью. Однако накопленный материал не позволяет однозначно решить вопрос о происхождении второго гумусового горизонта и он до сих пор остается предметом дискуссий.
Большинство исследователей считает второй гумусовый горизонт реликтом древнего почвообразования, результатом оподзо-ливания или осолодения (Иванова и Двинских, 1944), существовавших на этом месте в голоцене почв с развитым гумусовым профилем (черноземы, лугово-черноземные, лугово-болотные почвы и другие), под влиянием смены общеклиматических условий.
Некоторые авторы, не отрицая реликтового происхождения горизонта, выразили сомнение по поводу столь длительной сохранности древнего гумуса и предположили, что исходным про-
цессом формирования "реликтового" гумуса может быть гумификация торфа современных болотных почв при изменении их гидрологического режима (Глинка, 1923; Гаджиев, 1968) или временное преобладание дернового процесса над подзолистым при смене растительности (Тюлин и Россохина, 1967).
Другая, менее многочисленная группа исследователей считает второй гумусовый горизонт современным иллювиально-гуму-совым образованием (Горшенин и Сельская, 1929; Ильин, 1929; Тюрин, 1939; Кузнецов, 1948; Пономарева, 1956; Пономарева и Толчельников, 1968). Эта концепция наиболее полно сформулирована и аргументирована в работах В.В. Пономаревой (1956, 1968), где автор доказывает, что горизонт A2h образуется в настоящее время при постепенном насыщении и закреплении движущихся сверху неполно усредненных воднорастворимых гумино-вых кислот. Отмеченную ранее Д.А. Драницыным (1914) и И.М. Гаджиевым (1964), более яркую морфологическую выраженность горизонта у менее дренированных почв, она считает результатом внутрипочвенной склоновой миграции гумусовых растворов и их аккумуляцией в понижениях рельефа.
Веским аргументом в пользу реликтовой гипотезы считается абсолютный возраст гуминовых кислот (радиоуглеродные датировки). Анализ показал, что второй гумусовый горизонт дерново-подзолистых почв Западной Сибири старше верхнего на 4-6 тыс.лет (Добровольский и др., 1970; Толчельников, 1970). Но и здесь возникают сомнения. Во-первых, нижние горизонты всех без исключения естественных почв старше верхних (Арсланов и др.,1970; Герасимов, 1970). Во-вторых, некоторые почвоведы справедливо сомневаются в надежности измерения радиоуглеродным методом абсолютного возраста гуминовых кислот "живых" почв (Дюшофур, 1970; Ковда, 1969; Paul et al., 1964).
Отдельные авторы допускают возможность одновременного существования почв как с реликтовым A2h, так и с современным, образующимся в наши дни (Ковалев и Гаджиев, 1968; Наумов, 1960; Тюлин и Россохина, 1967). Такое допущение, видимо, следует считать правомерным ввиду отсутствия в настоящее время достаточно объективных аргументов, способных однозначно подтвердить или опровергнуть одну из гипотез.
В исследованных нами подзолистых и серых лесных почвах южной тайги Причулымья (Обь-Енисейское междуречье) второй гумусовый горизонт обнаруживается довольно часто в виде темной или черной полосы, расплывчатых темных пятен, ясно видимых на светлом фоне подзолистого горизонта. Почвы с A2h встречаются на различных элементах рельефа под темнохвойны-ми, лиственными и смешанными лесами, а также на типичных
здесь обширных гарях и "еланях" (полянах). Строгой приуроченности их к определенным элементам рельефа или типам растительности установить не удалось. На эту сложность указывают и другие авторы (Драницын,1914; Иванова и Двинских, 1944; Тю-лин и Россохина,1967 и др.).
Глубина залегания A2h колеблется в пределах 20-80 см от поверхности почвы. Степень его морфологической выраженности не всегда коррелирует с аналитическими данными. Иногда этот горизонт, хорошо выраженный морфологически, не выделяется по данным физико-химического анализа. И наоборот, слабо выраженный морфологически, он проявляется в гуматном составе органического вещества, на общем фульватном фоне.
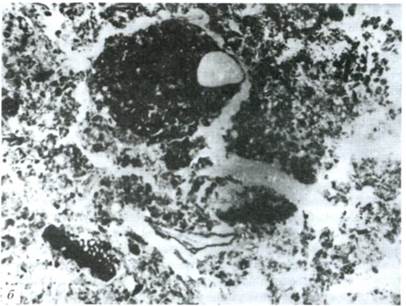
Микроморфологическое изучение этого горизонта выявило четыре формы морфологического проявления гумуса (рис. 6):
- микроортштейны (рис. 6а) - железистые стяжения или зерна минералов, покрытые черной глянцевой пленкой гумуса. Иногда между черной пленкой гумуса и минералом обнаруживается еще одна ярко-оранжевая глянцевая пленка. Размер 0,03-0,30 мм;
- микробрекчии (рис. 66) - округлые агрегаты, состоящие из мельчайших обломков различных минералов, сцементированных черной органической массой. Размер 0,4-0,8 мм;
- дробовины (рис. 6в) - черные шарики с глянцевой поверхностью и заметной на срезе волокнистой текстурой, подобной текстуре древесного угля, резко проявляют свойства гидрофобности. Размер шариков 0,7-1,0 мм;
- сыпь (рис. 6г) - тонкая черная пыль (0,01-0,03 мм), густо пропитывающая бурую колломорфную глину или заполняющая слепые внутриагрегатные поры.
Почти всегда в этом горизонте содержатся обугленные растительные остатки, сохранившие клеточное строение, а также мелкие и мельчайшие обломки древесного угля (0,01-0,02 мм), рассеянные по массе мелкозема.
В начальный период исследования почв Причулымья мы руководствовались гипотезой о реликтовой природе A2h (Керженцев, 1970). Затем, при более углубленном знакомстве с изменениями в пространстве характерных признаков этих почв, появились обстоятельства, заставившие усомниться в приложимости реликтовой гипотезы к почвам данного региона.
Во-первых, согласно реликтовой гипотезе, A2h должен раньше всего исчезнуть в дренированных почвах и дольше сохраняться в почвах понижений. Однако на водоразделах в Причулымье почвы с ярко выраженным A2h встречались гораздо чаще, чем вблизи естественных понижений.
|
|
Рис. 6. Формы гумуса в горизонте A2h. Увеличение 50.
а - микроортштейн (зерно минерала, покрытое пленкой гумуса); б - микробрекчии (обломки минералов, сцементированные гумусом)

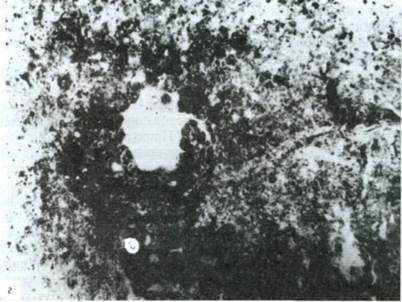
Рис. 6.
в - дробовина (с микроструктурой древесного угля); г - черная сыпь в колломорф-ной глине по краям пор и внутри агрегатов
Во-вторых, если A2h действительно реликт высокогумусного профиля, исчезающего под действием подзолистого процесса, то максимальной его сохранности надо ожидать в самых нижних горизонтах, куда еще не достиг процесс оподзоливания, тем более, по мнению И.П. Герасимова (1970) почва растет вверх. На самом деле темная полоса или серия темных пятен чаще всего выделяется на фоне элювиального подзолистого горизонта, где разрушаются даже минералы. Гумус слишком динамичная и неустойчивая система, чтобы сохранить все компоненты в течение 5-8 тыс. лет после коренного изменения породивших его условий. Кроме того, анализ качественного состава гумуса показал, что гумус A2h в разных почвах представлен разными фракциями гуминовых кислот, что указывает на отсутствие единого, общего для всех почв, реликтового предшественника.
С тех же позиций, приложимости к условиям Причулымья, была рассмотрена гипотеза В.В. Пономаревой (1956, 1968). В первой из упомянутых работ дана логичная и оригинальная трактовка процесса образования A2h, но без указания условий его возможного проявления, что лишает статью доказательности. В другой работе (Пономарева и Толчельников, 1968) авторы не столько аргументируют свою оригинальную гипотезу, сколько подвергают сомнению доказательства оппонентов. Кстати, Ю.С. Толчельников вскоре (1970) изменил свою точку зрения в пользу реликтовой гипотезы.
Мы попытались, насколько это возможно, определить те экологические условия, при которых образование второго гумусового горизонта A2h, согласно гипотезе В.В. Пономаревой наиболее вероятно, чтобы таким опосредованным способом подойти к объяснению его генезиса в почвах Причулымья.
Сразу же возник вопрос о принципиальной возможности образования гуматного гумуса в почвах южной тайги. Здесь встречаются дерново-глеевые почвы с темным гумусовым горизонтом. Но их формирование связано, как правило, с заболачиванием дерново-подзолистых или серых лесных почв. Этот гуматный гумус сформировался в анаэробных условиях.
Более вероятно для подзолистых почв южной тайги формирование гуматного гумуса после удаления лесного полога пожаром, вырубкой, инвазией энтомовредителей на больших площадях. В этом случае на поверхности почвы формируются гидротермические условия, идентичные условиям лесостепной зоны, которые стимулируют развитие дернового процесса с формированием темноцветного гумусового горизонта.
По оценкам Н.Г. Коломийца (1957), М.И. Куликова (1966), В.В. Фуряева (1966) только за период 1952-1957 гг. на террито-
рии Обь-Енисейского междуречья в Причулымье в результате инвазии сибирского шелкопряда погибло около 4 млн га высоко-бонитетных темнохвойных лесов. Такого масштаба инвазии в зоне южной тайги повторяются довольно регулярно.
Во время массового размножения миллионы гусениц сибирского шелкопряда уничтожают хвою и обрекают деревья на гибель. В результате крупные лесные массивы превращаются в кладбища мертвых деревьев, в пустыри, захламленные сухостоем и валежной древесиной. Площади отдельных очагов повреждения (шелкопрядников) колеблются от 500-700 га до 6-10 тыс. га, а в отдельных случаях даже до 70-150 тыс. га (Долгоунские гари).
Гибель древесного яруса сопровождается резким изменением всей экологической обстановки в шелкопряднике. В первую очередь изменяется фитоклимат территории. По наблюдениям М.И. Куликова (1966), летом среднемесячная температура приземного слоя воздуха в шелкопряднике повышается на 3-5 градусов, освещенность поверхности почвы возрастает в 2-3 раза по сравнению с неповрежденными участками леса.
Кроме этого, гусеницы сибирского шелкопряда после поедания хвои образуют значительную массу экскрементов, покрывающих поверхность почвы сплошным органо-минеральным слоем, который существенно меняет скорость и направленность биохимических процессов в почвах. По данным Н.Г. Коломийца (1957) за 3-4 недели активной жизни, гусеницы шелкопряда образуют на поверхности почвы слой экскрементов мощностью до 4-5 см, что составляет массу 16-30 т/га. Практически мгновенно формируется новый органический горизонт почвы. Каждая тонна экскрементов содержит в кедровниках 20,4 кг, в пихтачах -68,8 кг, в ельниках 124,0 кг кальция. Такая добавка стимулирует дерновый процесс в почвах и образование гуматного гумуса.
Столь глубокое изменение экологических условий сопровождается буйным развитием травянистой и кустарниковой растительности (Куликов, 1966; Фуряев, 1966). Увеличение количества тепла, аэрации и оснований активизирует микрофлору и фауну почвы, которая сверхактивно разлагает экстремально большую массу опада и отпада, в которой с годами начинает преобладать травянистая фитомасса. Эти изменения отражаются на ходе гумификации органических остатков, направляя процесс в сторону увеличения доли гуминовых кислот (Кауричев и др., 1970). На этом коренные преобразования профиля подзолистых почв не кончаются.
Через несколько лет участок тайги, поврежденный шелкопрядом, буквально превращается в склад сухой древесины, заросший густой травянисто-кустарниковой растительностью. Ог-
Таблица 3.2 Масштабы озоления фитомассы при выгорании шелкопрядника
| Вид горючего | Запас, | Сгорает | при пожаре | Зольность, | Выход золы, ц/га |
| материала | т/га | % | т/га | %% | |
| Древесина | 210,0 | 40-60 | 80-120 | 0,41 | 3,4-4,9 |
| Сучья | 24,0 | 30-60 | 7,2-14,4 | 2,62 | 18,9-37,3 |
| Опад | 9,5 | 70-80 | 6,7-7,6 | 3,06 | 20,5-23,3 |
| Подстилка | 32,0 | 10-30 | 3,2-9,6 | 5,96 | 19,1-57,2 |
| Ветошь трав | 5,5 | 30-50 | 1,7-2,8 | 3,21 | 5,5-9,0 |
| Всего | 281,0 | 98,8-154,4 | 68,9-130,2 |
ромные и равномерно распределенные по территории запасы сухой древесины (табл. 3.2) стимулируют возникновение частых и очень интенсивных пожаров, охватывающих обширные пространства. В Причулымье наблюдались пожары, при которых в течение 20-30 дн. выгорала площадь шелкопрядников 28-68 тыс. га (Фуряев, 1966).
Как видно из таблицы, во время пожара сгорает полностью 100-150 т/га органических остатков, в результате чего образуется 70-130 ц/га древесной золы. Налги измерения показали, что водная вытяжка древесной золы имеет сильнощелочную реакцию (рН 9-12), а щелочь, как известно, является хорошим экстраген-том почвенного гумуса и вполне может содействовать его транзиту из верхнего горизонта в нижележащие. Высокая кислотность на контакте горизонтов А2 и В,, способствует осаждению гумино-вых кислот именно в этом месте почвенного профиля.
Мы провели эксперимент по оценке возможного изменения рН почвы в результате пожара. На бумажных фильтрах в стеклянных воронках помещали навески 10 г из смеси фракций в различных сочетаниях: подзолистой почвы (гор. А^), озоленной и обугленной массы опада пихтача разнотравного. Навески промывали многократно дистиллированной водой. После каждой промывки измеряли рН фильтратов.
Результаты измерений, приведенные в табл. 3.3 показали, что даже после 15 промывок фильтраты с участием озоленных или обугленных остатков оставались щелочными, а фильтрат подзолистой почвы - кислым.
Фильтраты от промывки смеси золы с почвой и особенно золы с углем имели бурую окраску, что указывает на совместное передвижение в растворах оснований и гумусовых веществ. Сле-
Таблица 3.3
Дата добавления: 2020-10-01; просмотров: 738;
Поиск по сайту
Узнать еще
- II Классификация САSЕ-средств
- II. Затраты (времени, физических сил, финансовых средств и т.д.)
- II. Классификация документов
- II.4. Классификация нефтей и газов по их химическим и физическим свойствам
- III. Классификация методов воспитания.
- III.1.3. ПРИЧИНЫ НАРУШЕНИЙ СЛУХА. ПСИХОЛОГО-ПЕДАГОГИЧЕСКАЯ КЛАССИФИКАЦИЯ НАРУШЕНИЙ СЛУХОВОЙ ФУНКЦИИ У ДЕТЕЙ
- III.2.3. ПРИЧИНЫ НАРУШЕНИЙ ЗРЕНИЯ. КЛАССИФИКАЦИЯ НАРУШЕНИЙ ЗРИТЕЛЬНОЙ ФУНКЦИИ У ДЕТЕЙ
- VI.2. Классификация месторождений нефти и газа

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
