Биоразнообразие и динамика экосистем: информационные технологии и моделирование. Отв. ред. В.К. Шумный и др. Новосибирск. Изд-во СО РАН. 2006. 648 с. 1 глава
ПОПУЛЯЦИЯ - 1. Совокупность особей одного вида, занимающих определенную территорию, свободно скрещивающихся друг с другом, имеющих общее происхождение, генетическую основу и в той или иной степени изолированная от других популяций данного вида.
П. - элементарная эволюционная структура. Обладает рядом признаков, отсутствующих у отдельных ее членов. Контакты между особями внутри одной П. происходят чаще, чем между особями разных П.
П. способна к развитию, в ней протекают простейшие эволюционные процессы.
ЛокальнаяП. -географически отделенная в той или иной степени от др. группа особей одного вида.
П. – Совокупность особей одного вида, длительно населяющих определенное пространство, внутри которого практически осуществляется та или иная степень панмиксии.
П. отделена от соседних аналогичных совокупностей той или иной степенью давления различных форм изоляции и имеет сложную структуру по полу, возрасту, пространственным и близкородственным объединениям особей.
П. – элементарная единица эволюционного процесса и форма существования вида. Каждая П. способна (теоретически – неограниченно долго) развиваться при подходящих условиях окружающей ее среды.
Основными характеристиками П. являются: численность, плотность, рождаемость, смертность, прирост особей, темп роста и др.
П. имеет определенную структуру: возрастную (соотношение особей разного возраста), сексуальную (соотношение полов) и пространственную (колонии, семьи, стаи). Так, возрастная структура П. является ее важной характеристикой, влияющей на рождаемость и смертность. Соотношение разных возрастных групп в П. определяет ее способность к размножению в данный момент, причем обычно в быстрорастущих П. значительную долю составляют молодые особи. П. реагируют на изменения внешней среды путем колебания численности, соотношения полов, возрастной структуры и т. д. Численность особей в П. колеблется в значительных пределах, однако она не может быть ниже некоторых пределов. Сокращение численности за эти пределы может привести к вымиранию П. Полагают, что при численности П. меньше нескольких сотен особей какие-либо случайные причины (пожар, наводнение, обильные снегопады, сильные морозы и т.п.) могут сократить численность настолько, что оставшиеся особи не смогут встретиться и оставить потомство. В соответствии с величиной популяционного ареала и численностью особей находится плотность особей в П. Соотношение половозрелых и неполовозрелых особей в П. различно и зависит от продолжительности жизни, времени наступления половой зрелости, интенсивности размножения. Генетический механизм определения пола обеспечивает расщепление потомства по полу в соотношении 1:1. В силу разной жизнеспособности мужских и женских организмов это первичное соотношение порой заметно отличается от вторичного (при родах – у млекопитающих) и тем более от третичного (характерного для взрослых особей). Например, в П. человека вторичное соотношение полов 100 девочек:106 мальчиков; к 16-18 годам это соотношение выравнивается и становится 1:1 (100 девочек:100 мальчиков), к 50 годам – 100 женщин:85 мужчин, а к 80 годам соотношение по полу становится 2:1 (100 женщин:50 мужчин).
Размеры П. (пространственные и по числу особей) подвержены постоянным колебаниям. (Периодические колебания численности П. называются волнами жизни, или популяционными волнами.) Причины этих колебаний различны и в общей форме сводятся к влиянию биотических и абиотических факторов (влага, микроорганизмы, вызывающие заболевания, запас пищи, свет, температура, конкуренты, стихийные, бедствия и т.п.). Масштабы таких измерений могут быть весьма значительными. Так, например, на одном небольшом острове у юго-западного побережья Англии обитает в диком виде П. кроликов. В период максимальной численности (осенью благоприятного по кормовым условиям года) общее число кроликов достигало 10 тысяч особей, а однажды (после холодной и малокормной зимы) число их сократилось до 100 особей. С изменением числа особей в П. изменяется их плотность, т.е. число особей на единицу площади. (Определенная плотность П. поддерживается за счет конкуренции.) В отдельные годы резко увеличивается плотность П. насекомых-вредителей, кровососов, грызунов, сорных и др. растений. Верх, предел плотности П. определяется количеством самого дефицитного ресурса (воды или питательных элементов для растений, растительной пищи для травоядных животных, количеством жертв для хищников). Устойчивость П. поддерживается исторически сложившимися способами самовоспроизведения благодаря смене поколений и способности к саморегуляции путем изменения своей структуры. Например, в П. жука мучного хрущака при плотности П., превышающей оптимальную, внутривидовая конкуренция превращается в каннибализм – самцы поедают яйца, откладываемые самками, т.е. осуществляется изменение возрастной структуры П. и снижается ее плотность. У некоторых видов увеличение плотности П. вызывает резкое сокращение или вообще временную утрату способности давать потомство. У видов растений, не имеющих специальных приспособлений для распространения семян на большое расстояние, состояние перенаселенности (высокой плотности) возникает достаточно часто. В этих случаях, чтобы предотвратить очень резкую внутривидовую конкуренцию, уменьшается размер растений. Например, растения мари белой в зависимости от плотности П. могут отличаться по размерам в сотни раз. У этого вида развиваются практически все растения из всхожих семян. Однако меньшие по величине растения дадут и меньше семян. В результате, чем выше плотность П., тем меньше образуется семян, а в итоге плотность П. остается либо на прежнем уровне, либо снижается. В П. люцерны при плотности выше оптимальной растения не цветут и не плодоносят, но при этом происходит интенсивное накопление запасных питательных веществ в корневой шейке, которые используются для цветения и плодоношения, когда восстановится оптимальная плотность растений на единицу площади. Т. обр., регуляция численности и плотности особей в П. регулируется абиотическими и биотическими факторами, в т. ч. конкуренцией, а также внутрипопуляционными механизмами.
Судьба вида в биоценозе.Виды, составляющие каждый конкретный биоценоз, различаются своими требованиями к окружающей среде. Поэтому биологические особенности вида лежат в основе его экологической индивидуальности в структуре биоценоза. Своеобразие видов проявляется в различных требованиях к вещественным и энергетическим условиям среды, необходимым для роста и развития, в темпах размножения как основной функции, обеспечивающей положение вида в сообществе.
Попытки объяснить судьбу вида в биоценозе привели к формированию концепций стратегий вида и концепции экологической ниши.
В начале XX в. Л.Г. Раменский сформулировал концепцию экологической индивидуальности вида, основанную на идее конкурентных отношений за вещественные и энергетические ресурсы в каждом конкретном месте земной поверхности.
Ему удалось выделить три основных типа стратегии вида в биоценозе.
1. Виды, обладающие высокой конкурентоспособностью и энергией жизнедеятельности, полнотой использования ресурсов среды, названные им виолентами (более образно — «львами»). Это абсолютные конкуренты, способные вступать в борьбу за жизненный успех со всеми другими видами. Число их, видимо, невелико.
2. Виды, обладающие низкой конкурентоспособностью, но зато быстро осваивающие новые территории. Они получили название эксплерентов (или «шакалов»). Это пионерные виды. Они быстро заселяют свободные территории, но так же быстро и уходят с них. Число их также невелико.
3. Виды, обладающие большой выносливостью в различных условиях, получили название патиентов («верблюдов»). Это основная масса видов. Различные группы патиентов могут обладать чертами как виолентов, так и эксплерентов.
Т. А. Работнов, разрабатывая данную концепцию, пришел к выводу, что виолентность, патиентность, эксплерентность присущи не только виду в целом, но проявляются и на уровне отдельных популяций и особей.
Э. Пианка предложил другой подход к выделению стратегий вида, основанный на учете доли энергии, используемой на размножение, от общих затрат на жизнедеятельность. В рамках этой концептуальной основы появились представленияо k- и r-стратегиях. Общие характеристики видов, обладающих такими стратегиями, приведены в табл. 5.5. Дальнейшая разработка этой концепции привела к выделению различных промежуточных вариантов с различным сочетанием свойств обеих стратегий.
В общем плане r-стратегия характеризуется большим энергетическим вкладом в производство потомства, а k-стратегия — затратами на поддержание жизненной активности, т. е. на рост и поддержание во взрослом состоянии.
Другая концепция основана на понятии экологической ниши. Термин «ниша» впервые применил для обозначения самой малой единицы распространения вида Р. Джонсон в 1910 г. Через 17 лет, в 1927 г., Ч. Элтон отметил функциональный аспект этого термина и определил его как место вида в структуре биоценоза. Идея экспериментально исследована Г. Ф. Гаузе, Т. Парком и др., что позволило сформулировать эмпирическое обобщение: «Полные конкуренты не могут существовать бесконечно». Из обобщения следует, что если два вида существуют вместе, то между ними должно быть какое-то биологическое различие, обусловливающее различные экологические ниши обоих видов.
Таблица
Общая характеристика видов, обладающих разными стратегиями
| r-стратегия | k-стратегия |
| Размножаются быстро | Размножаются медленно |
| Скорость размножения не зависит от плотности | Скорость размножения зависит от плотности |
| Потомков много. Отсутствует забота о потомках | Потомков немного. Забота о потомстве |
| Вид не всегда устойчив на данной территории | Вид устойчив |
| Расселяются широко. У животных может мигрировать каждое поколение | Расселяются медленно |
| Малые размеры особей | Крупные размеры особей |
| Небольшая продолжительность жизни отдельной особи | Большая продолжительность жизниотдельной особи |
| Местообитание сохраняют недолго | Местообитание устойчиво |
| Слабые конкуренты | Сильные конкуренты |
| Слабые защитные приспособления | Хорошие защитные механизмы |
| Не становятся доминантами | Могут становиться доминантами |
| Слабая специализация | Высокая специализация |
Д. Хатчисон (1958) считал, что нишу следует определить в рамках всего диапазона физических, химических, биотических переменных среды. В идеале каждую такую переменную можно рассматривать как некий градиент, на котором у каждого вида имеется свой диапазон активности.
Ю. Одум (1971) настаивает на придании нише функционального аспекта («профессии» вида в биоценозе), оставив остальные аспекты в рамках понятия «местообитание».
В общем плане экологическая ниша — это процесс дифференцировки пространственно-временных отношений между видами, составляющими конкретный биоценоз.
Теоретически, при исключении всяких давлений извне, каждый вид способен освоить всю совокупность условий в конкретном месте. Это его потенциальная, или фундаментальная, ниша. Наличие других видов не позволяет ее реализовать в полной мере. Поэтому реальная экологическая ниша всегда будет частью фундаментальной. В ходе процесса дифференцировки пространства, занятого биоценозом, экологические ниши видов могут проявляться как независимые, с разной степенью перекрытия, и ниша одного вида может находиться внутри ниши другого.
Концепции экологической индивидуальности вида и экологической ниши, рассматривая разные стороны судьбы вида в биоценозе, дополняют друг друга. Остается неразработанным вопрос о роли стратегий видов в формировании структуры биоценоза: как виды с разной стратегией формируют ниши, чем отличаются экологические ниши видов разных стратегий, за счет особей каких стратегий решаются те или иные вопросы взаимоотношений с окружающим миром.
Устойчивость биоценоза является характеристикой, наиболее интересной для оценки его связей с окружающей средой, т. к. она выступает как высший интегральный показатель всех процессов, протекающих внутри биоценоза.
До 70-х гг. XX в. считалось, что устойчивость сообщества определяется видовым разнообразием: чем больше видов, тем устойчивей биоценоз к различным внешним воздействиям. Теоретическое осмысление этой идеи, разработанные разнообразные математические модели, экспериментальные исследования и наблюдения в природе привели к формированию трех основных взглядов на данную проблему:
• устойчивость биоценоза повышается с увеличением видового разнообразия;
• устойчивость биоценоза снижается с увеличением видового разнообразия;
• устойчивость всех биоценозов Земли примерно одинакова, т. е. является некоторой константой.
Каждая из этих точек зрения имеет своих сторонников и противников и, что особенно важно отметить, определенную систему доказательств: от теоретических посылок, математических моделей до наблюдаемых явлений в природе. М. Бигон, проведя достаточно большой анализ данной проблемы, заканчивает его следующими словами:
...Ясно одно: такой вещи, как устойчивость сообщества, вообще попросту не существует. Этот параметр зависит от исследуемого компонента системы и характера нарушения.
Рассмотрение биоценоза как биотической компоненты экосистемы позволяет несколько прояснить этот вопрос. Поскольку экосистема есть мыслительная конструкция, выделяемая каждый раз для решения конкретной задачи, то и характеристика ее устойчивости определяется также в рамках задачи.
Сущность биоразнообразия.Реализация генетической информации происходит на клеточном уровне организации жизни. Под действием разных факторов могут происходить мутации – наследуемые изменения генетического материала. В результате мутаций возникают различные структурно-функциональные варианты генов – аллели (аллеломорфы), которые часто приводят к изменению проявления тех или иных признаков. Совокупность генов (включая различные аллели у диплоидных и полиплоидных организмов) каждой особи называют генотипом. Генотип контролирует развитие, организацию (морфологию и анатомию) и функционирование (физиологию, поведение) особи, т.е. набор всех ее свойств и признаков, обычно обозначаемых как фенотип, поэтому в ряде случаев его определяют как совокупность генов, имеющих фенотипическое проявление.
При половом размножении (или его аналогов у прокариот и некоторых эвкариот) происходит рекомбинация – перераспределение генетического материала родителей у потомства. Таким образом, в результате мутационного и рекомбинационного процессов создается генетическое разнообразие особей вида.
Большая часть молекул ДНК эвкариотических организмов находится в клеточном ядре в составе хромосом. Под световым микроскопом во время деления клетки их можно сосчитать, идентифицировать по длине и ряду других параметров. Характерный для данной особи набор хромосом называется кариотипом. Внутривидовая изменчивость кариотипов обычно незначительна, поэтому их сравнительный анализ используют для установления видовой принадлежности и родственных отношений между таксонами. Вместе с тем в некоторых группах (особенно при анализе политенных хромосом) удается выявить и межпопуляционную изменчивость кариотипов.
У эвкариот часть ДНК находится в митохондриях и хлоропластах. Эта так называемая внеядерная ДНК отличается по ряду характеристик от ядерной и, как правило, наследуется только по материнской линии. В последние годы сравнение первичной структуры фрагментов как ядерной, так и внеядерной ДНК часто используют для характеристики биоразнообразия и эволюционных отношений между разными группами живых существ.
Особи с разными генотипами могут иметь практически одинаковый фенотип и, наоборот, особи с одинаковыми генотипами могут существенно различаться фенотипически, поскольку на реализацию генотипа отпечаток накладывают конкретные условия существования, т.е. факторы окружающей среды. Такие изменения определяются нормой реакции –возможным размахом фенотипической изменчивости без изменения генотипа. Норма реакции определяет пределы возможностей существования особи в меняющейся окружающей среде – ее экологическую амплитуду, степень приспособленности (адаптированности) каждой конкретной особи к существованию в экосистеме.
Для характеристики биоразнообразия на данном уровне организации существен также учет характера онтогенеза, т.е. индивидуального развития, – процесса преобразования особи от зарождения (образования зиготы) до смерти одной особи или ряда поколений вегетативно возникших особей от диаспоры до естественной смерти в результате старения. В ходе онтогенеза нередко значительно меняется разворачивание информации, содержащейся в генотипе. Часто прослеживаются стадии индивидуального развития, причем переход из одной стадии в другую может быть связан с метаморфозом, т.е. с кардинальной перестройкой организма (например, у насекомых с полным превращением). В результате нередко меняются фенотип, экологическая амплитуда и даже жизненная форма.
Из особей состоит каждая популяция, а в итоге и вид в целом. Основными понятиями этого уровня являются популяция и вид. По А.В. Яблокову [1987], популяция– это минимальная самовоспроизводящаяся группа особей одного вида, на протяжении эволюционно длительного времени населяющая определенный участок земной поверхности, образующая самостоятельную генетическую систему и формирующая собственное гиперпространство. Реально это определение относится к локальной популяции, или дему. Каждая такая популяция отличается определенной структурированностью. Нередко популяциями называют приуроченные к какой-то территории или акватории совокупности демов.
Фактически это отражает сложную иерархическую организованность популяционной системы вида, включающей всю совокупность взаимосвязанных и взаимодействующих (хотя бы в прошлом) популяций данного вида. Естественно, на каждую популяцию и популяционную систему в целом воздействуют разные факторы окружающей среды. Следует также отметить, что нередко происходит выселение групп особей за пределы основного ареала вида или популяции. В результате образуются временные поселения, которые могут существовать даже на протяжении нескольких поколений, но которые не приводят к формированию настоящих локальных популяций.
Определение понятия вид связано со значительными сложностями (особенно в плане описания биоразнообразия). Однако несомненно, что вид –это более или менее целостная совокупность популяций, изолированная (в том числе с эволюционной точки зрения) от других таких же совокупностей и характеризующаяся набором определенных признаков (в том числе областью распространения – ареалом) Нередко разделяют представления о морфологическом виде, включающем особи с определенным набором отличий от других морфологических видов, и о биологическом виде, объединяющем совокупность популяций, между которыми возможен или существует обмен генетической информацией. Первое представление используется главным образом систематиками, а второе – в эволюционно-генетических исследованиях. Вид можно рассматривать и как открытую кибернетическую систему, регуляция которой основана на положительных и отрицательных обратных связях; такая система взаимодействует с окружающей средой благодаря наличию входящих и выходящих каналов связи, контролируемых самой системой.
Для поддержания целостности биологического вида важны миграции, т.е. перемещения в широком смысле особей из одного места в другое, а обособленность таких видов друг от друга в значительной степени обеспечивается изоляцией благодаря наличию разного рода барьеров (физико-географических, экологических, морфологических, физиологических и т.п.), препятствующих свободному скрещиванию. Это особенно существенно для групп, размножающихся половым путем. Вместе с тем недавно показано, что географические барьеры играют свою роль и в дифференциации поселений океанических прокариот, для которых обычно постулируется неограниченная способность к расселению.
___________________________
6. Ареал вида. Факторы, влияющие на ареалы древесных растений.
Древесные растения, как и другие группы растений, распространены на земной поверхности не беспорядочно, они приурочены к определенным территориям. Одни виды широко встречаются на огромных территориях, выходящих иногда за пределы целого континента, тогда как другие произрастают на незначительных участках. Вся область обитания вида, рода или другой таксономической категории называется его ареалом.
Если на географическую карту нанести все точки нахождения вида, то получим точечное изображение его ареала (точечный ареал). Если теперь соединить линией все точки крайних местонахождений вида, то будет очерчена граница ареала, выявлена его площадь, внутри которой развивается и процветает данный вид. Территория, ограниченная этой линией, и является площадью его географического ареала.
Можно не обводить границы ареала сплошной линией, а ограничиться точками географических пунктов. Точечное изображение ареала отражает не только географические границы объекта, но и его распределение по территории. Обычно средние части площади ареала густо покрыты точками — это означает, что здесь растение встречается часто и находится в оптимальных условиях обитания. Ближе к границе ареала пунктов нахождения меньше — растение все реже находит для себя благоприятные условия существования. Чаще всего вид встречается рассеянно среди других видов растений или даже приурочен только к определенным орографическим или эдафическим условиям: например, сосна обыкновенная растет по песчаным отложениям вдоль рек и по берегам озер, тогда как водораздельные пространства с суглинистыми и глинистыми почвами на значительных площадях заняты елью и другими породами.
По форме на карте ареалы растений делят на три типа: сплошные, разорванные и ленточные.
В сплошном ареале вид растения занимает территорию без каких-либо больших перерывов, осуществляется постоянный обмен генами между популяциями вида.
Ареал называется разорванным, если территория, занятая видом, распадается на две или более обособленные части, между которыми обмен генетическим материалом невозможен. Например, липа мелколистная из Европы в Западной Сибири распространяется на восток до Тюмени. Затем следует разрыв на много сотен километров, и снова липа появляется уже в предгорьях Саян. Другой пример — ареал кедра сибирского, произрастающего, в основном, в Сибири. В европейской части России его нет. Затем он снова появляется в Карпатах и Альпах.
Ленточные ареалы обычно имеют виды, приуроченные к современным и древним речным долинам (тополь черный, ольха черная, ивы). В ленточные могут переходить сплошные и разорванные ареалы на северных и южных границах. Такую форму имеют, например, ареалы дуба черешчатого и сосны обыкновенной в лесостепи и степи.
С течением времени площадь и границы ареала не остаются неизменными. Под влиянием разных причин они могут заметно изменяться как в сторону уменьшения, так и расширения. Часто сокращение ареала вызывается деятельностью человека: вы рубка лесов, распашка лесных площадей, изменение водного режима значительных лесных территорий при сооружении гидротехнических станций и т.д. Могут измениться природные условия и по естественным причинам (изменение климата, заболачивание или осушение территории). В этом случае данный вид не сможет в дальнейшем существовать в прежних границах ареала. Возможны и противоположные изменения природных условий, которые вызывают расширение ареала вида. Такой ареал принято считать прогрессивным. При сужении ареала в связи с ухудшением природных условий ареал считается регрессивным.
Если растения обитают на довольно ограниченной территории и нигде в других районах не встречаются, то такие виды называют эндемичными или эндемами. Например, ель восточная на Кавказе микробиота в Приморье.
Очевидно, что условия существования вида оптимальны в центральной части ареала, а менее благоприятны — на его окраинах. Иногда растение за пределами своего ареала встречает лучшие условия для своего развития и быстро там акклиматизируется.
Знание ареала каждого вида растений имеет большое практическое значение. Сопоставление всех факторов внешней среды, обеспечивающих существование данного вида в определенных его границах, позволяет судить о возможности выращивания его в других районах.
_______________________
http://big-archive.ru/geography/general_geography/50.php
Ареалы видов растений.Для выяснения географических закономерностей распространения растительных видов используют метод ареалов. Метод применим также для изучения распространения растительных сообществ.
Ареалом называют часть поверхности земли, в пределах которой встречаются определенные растения (например, дуб, лиственница даурская, финиковая пальма). Поскольку экологические условия в пределах ареала меняются, степень заселенности ареала будет различной. В зависимости от распространения вида ареалы подразделяют по форме на сплошные, прерывистые и ленточные.
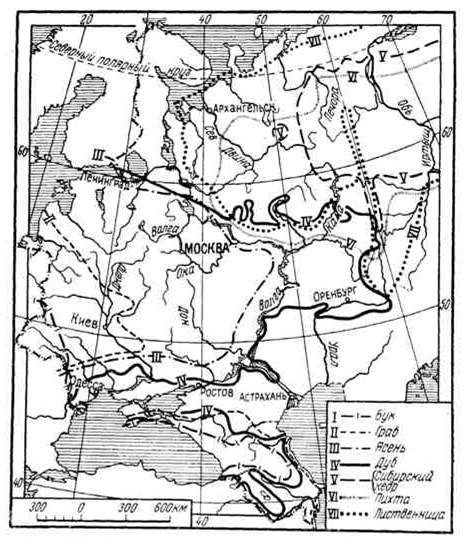
Рис. Ареалы древесных видов в Европейской части СССР
Сплошным ареалом считают такой, в пределах которого нет непреодолимых препятствий для расселения растительного вида (рис.).
Прерывистые (разорванные) ареалы представляют собой участки, населенные видом обособленно. Примером может служить ареал кислицы, распространенной в Северной Америке, Европе, Восточной Сибири, в Гималаях и горах Западного Китая, ареал дуба, высокогорные альпийские виды (камнеломка, толокнянка и др.).
Ленточные ареалы приурочены к поймам, террасам скалистым берегам речных долин, побережьям (пойменные луга, сосновые боры на террасах, мангровые леса и др.).
Причины разрыва ареалов разнообразны. Например, изменение климатических условий, результат миграции растений, результат деятельности человека. Но главной причиной, объясняющей площадь распространения ареалов и их форму, является история развития суши и ее флористических областей.
__________________________
7. Половая и гибридогенная формы изменчивости древесных растений.
Склонность к изменчивости представляет собой такое же свойство природы, как и самые фундаментальные физические законы. Она есть следствие толо факта, что условия, сопровождающие появление, казалось бы, совершенно однородных тел, не могут быть идеально одинаковыми по причине влияния на этот процесс множества воздействий. Можно говорить о стремлении природы к изменчивости, лишь сдерживаемой теми или иными факторами. Наследственность немыслима без изменчивости. Весь процесс эволюции жизни на земле есть не что иное, как проявление в природе стремления к изменчивости (Хохряков, 1975).
Изменчивость не объясняется всецело внешними влияниями. Внутренние факторы, заложенные в самом организме, не лишены при этом какого-либо значения, так как для изменчивости, являющейся своего рода ответом организма на внешние воздействия среды, организм, конечно, значительно важнее, чем среда (Филипченко, 1978).
При определении половой изменчивости у двудомных видов древесных растений учитывается соотношение мужских и женских особей. У двудомных культур (облепиха, тополь, ива и др.) сорта и формы разделяются на четыре типа: 1) женские особи (мужские почки отсутствуют). 2) мужские особи (мужские почки по всей кроне). 3) особи с яным преобладанием мужских почек, женские – единичны. 4) особи с яным преобладанием женских почек, мужские – единичны.
Для изучения хронографической изменчивости плодов подбирается возрастной ряд насаждений, произрастающий в одном природном районе в одинаковых условиях местообитания. Элементы возрастного ряда, например, из облепихи должны различаться между собою на 2-3 года, так как срок стабильного плодоношения этой культуры ограничен до 10-15 лет.
__________________________
8. Экологическая и географическая форма изменчивости древесных растений.
Географическая изменчивость связана с внутривидовыми таксонами разного геграфического происхождения, которые создаются посевом семян. Минимальное количество – по 30 сеянцев каждой географической расы.
________________________
Дата добавления: 2020-08-31; просмотров: 1097;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
