Репродуктивное поведение н физиология
Сезонные климатические изменения (температуры и осадков) оказывают мощное влияние на успех размножения многих видов. У птиц наличие корма для птенцов, по-видимому, главный определяющий фактор такого успеха (Lack, 1968). В средних и северных широтах размножение птиц связано с временем года; обычно они откладывают яйца весной, что дает возможность птенцам достаточно созреть, чтобы противостоять зимним условиям или выдержать долгий миграционный перелет. Так, например, песочники, размножающиеся в арктических областях, строят гнезда и высиживают птенцов весной, ког-
да земля еще покрыта снегом. Обычно птенцы вылупляются, когда снег тает и в изобилии появляются насекомые, которые служат для них кормом (West, Norton, 1975).
Репродуктивная физиология у сезонно размножающихся животных привязана к годичному циклу изменений среды таким образом, что появление детенышей предваряет пик изобилия корма или неблагоприятные климатические условия. Это осуществляется двумя способами. Во-первых, изменения внешней температуры, продолжительности светового дня или других факторов среды вызывают физиологические изменения в определенное время года. Во-вторых, сезонные физиологические изменения запрограммированы посредством эндогенных цирканнуальных часов.
Самое регулярное и предсказуемое изменение среды связано с продолжительностью светового дня. У прозрачных организмов свет может прямо действовать на гонады, приводя их в репродуктивное состояние в надлежащее время (Scharrer, 1964). Некоторые другие животные имеют прозрачное «окно», которое пропускает свет в головной мозг. Нейросекреторные клетки превращают световые стимулы в химические сигналы. У ряда млекопитающих эпифиз, расположенный на дорсальной поверхности головного мозга, может действовать как преобразователь света (Wurtmann et al., 1968). Но свет влияет на размножение млекопитающих, главным образом действуя через сетчатку на гипоталамус, как показывает схема Шаррера (рис. 16.7).
Регуляция физиологии размножения состоит в сложном взаимодействии ряда гормонов (рис. 16.8). У большинства позвоночных факторы среды стимулируют выработку гонадотропных гормонов гипофизом. Эти гормоны стимулируют рост и активность семенников и яичников, которые в свою очередь производят характерные половые гормоны. К концу сезона размножения активность гипофиза снижается, гонады также становятся неактивными и репродуктивное поведение затухает. Кроме сезонных циклов половой активности у многих млекопитающих
имеется гораздо более короткий цикл - эструс, или «течка». У одних животных, например у рыжей лисицы (Vulpes vulpes), течка бывает только раз в году; у других, например у домашних собак, - два раза; у
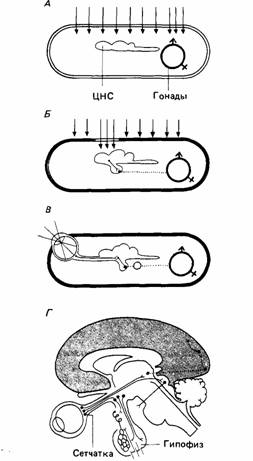
| Рис. 16.7. Влияние света на гонады по Шарреру (Scharrer, 1962.) А. У прозрачных животных свет может оказывать прямое действие на внутренние органы. Б. Некоторые непрозрачные животные обладают прозрачным «окном», пропускающим свет к фоточувсгвительным областям головного мозга, который затем стимулирует гонады с помощью гормонов, выделяемых гипофизом. В. Система, в которой свет действует на сетчатку, посылающую нервные сигналы гипоталамусу. Эта область мозга стимулирует выделение гипофизом гонадотронных гормонов. Г. Система В применительно к головному мозгу человека. (По Bligh, 1976.) |
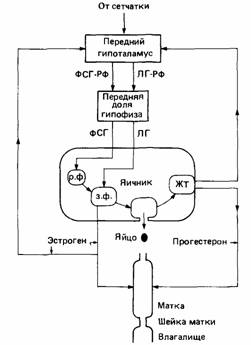
| Рис. 16.8. Главные гормональные пути, участвующие в работе яичника у млекопитающего. ФСГ-РФ -фактор, стимулирующий высвобождение фолликулостимулирующего гормона (ФСГ); ЛГ-РФ фактор, высвобождающий лютеинизирующий гормон (ЛГ); р. ф.- растущий фолликул; з. ф.- зрелый фолликул; ЖТ- желтое тело. (По Bligh, 1976.) |
третьих - чаще. У птиц число выводков в одном сезоне может зависеть от количества корма (Lack, 1968).
На характер размножения многих видов влияют как фотопериодические факторы, так и эндогенные цирканнуальные часы. Предполагается (Follett, 1973), что эти два механизма могут взаимодействовать. На виды, обитающие в высоких широтах, обычно сильнее всего влияет фотопериодичность. В низких (экваториальных) широтах годичных фотопериодических изменений меньше, но тем не менее годичный репродуктивный цикл может иметь свои преимущества. Например, у животных, обитающих в безводной пустыне, начало процесса размножения может зависеть от выпадения дождя (Marshall, 1970). Некоторые экваториальные виды обладают выраженным ритмом размножения, который, по-видимому, не связан с сезонными изменениями. Ряд морских птиц, в том числе коричневая олуша (Sula leucogaster), темная крачка (Sterna fuscata) итонкоклювая крачка (Anaus tenuirostris), размножаются каждые 8-10 месяцев. Постепенное расхождение между такими циклами размножения и годичным циклом говорит о том, что в экваториальных условиях ни одно время года не является предпочтительным. Почему же в таком случае птицы не размножаются непрерывно? Возможно, это потребовало бы слишком много энергии, и оптимальным вариантом становится периодический отдых, во время которого может происходить линька.
Годичный цикл размножения сходен с акклиматизацией тем, что при нем также происходят медленные физиологические процессы, коренным образом меняющие физиологическое состояние животного. Репродуктивная деятельность, включающая в себя защиту территории, ухаживание, спаривание и заботу о потомстве, требует от животного дополнительной энергии и физиологических затрат, которые должен обслуживать весь его организм. Если в данный год эти затраты так велики, что животное не в состоянии сохранить физиологическую стабильность, оно вынуждено отказаться от размножения до более благоприятного года.
Зимняя спячка
В частях света со сменой времен года животным иногда приходится приспосабливаться к длительным периодам неблагоприятной погоды. Некоторые из них избегают таких условий путем миграции, а другие способны пережить их, впадая в продолжительный сон, который при высоких температурах называется эстивацией (летней спячкой), а при низких - гибернацией (зимней спячкой). Некоторые пустынные грызуны, например суслики (Citellus), летом впадают в оцепенение, при котором температура тела падает и происходит общее снижение физиологической активности. Это значит, что затраты энергии уменьшаются и животное может долго жить, не питаясь. Потери воды сокра-
щаются вследствие меньшего потребления пищи, чему способствует уход в нору. Некоторые грызуны запасают корм в своих норах и сохраняют в них высокую влажность, закупоривая вход (Schmidt-Nielsen, 1964). Летняя спячка наиболее распространена среди обитателей пустыни, но не ограничивается ими. Так, например, многие европейские виды дождевых червей летом спят. Каждое животное вырывает небольшую нору глубоко в земле и сворачивается в ней в клубок. Наступление такой летней спячки связано с низкой влажностью, и ее можно предотвратить содержанием червей во влажной атмосфере.
В зимнюю спячку впадают многие виды в северных широтах, что позволяет им избегать зимних условий, требующих очень больших энергетических затрат. Подлинная зимняя спячка отличается от частичной спячки, в которую впадают европейский бурый (Ursus arctas) и американский черный (Ursus americanus) медведи. При частичной спячке температура тела медведя может упасть примерно от 38° до 30°С, хотя температуры тела ниже 15°С детальны. При истинной зимней спячке температура тела может упасть даже до 2°С. Истинная зимняя спячка характерна для мелких млекопитающих, хотя сходные с ней виды оцепенения бывают у енотов, барсуков и некоторых птиц. У многих видов колибри во время оцепенения температура тела снижается до температуры среды, хотя температуры ниже 8°С детальны. У птиц оцепенение длится обычно всего несколько часов, а сезонная спячка известна только в одном их семействе - у козодоев (Caprimulgidae).
Период спячки, называемый диапаузой, как реакция на неблагоприятные климатические условия бывает также у насекомых. Диапауза часто связана с определенной стадией жизненного цикла. Насекомые, избегающие замерзания с помощью переохлаждения, могут пережить сильный холод в диапаузе, при которой температура замерзания жидкостей тела намного ниже 0°С. Глубина переохлаждения может зависеть от степени акклиматизации, которая постепенно меняет химический состав жидкостей тела. Канадская оса Bracon cephi может снизить точку их замерзания до — 46° С, повысив концентрацию глицерола в гемолимфе.
Истинная зимняя спячка наступает только у мелких млекопитающих, которые охлаждаются быстрее крупных из-за относительно большей поверхности тела. Они также быстрее согреваются благодаря своей малой теплоемкости. Зимняя спячка характеризуется сноподобным состоянием с замедленными дыханием и сердцебиением. При этом животные часто выбирают особое место и принимают позу, в которой обычно спят. При зимней спячке температура тела и затраты энергии падают ниже уровня, характерного для нормального сна. У таких животных часто откладывается специальный бурый жир. Его главная функция состоит в образовании тепла, а не энергии для обменных процессов, для которых мобилизуются резервы обыкновенного жира. Особенно важную роль бурый жир играет в период пробуждения от спячки, когда температура тела должна быстро повыситься.
Некоторые млекопитающие, в том числе золотистый суслик (Citellus lateralis) и североамериканский лесной сурок (Marmota топах), обладают выраженным цирканнуальным ритмом, лежащим в основе их сезонной зимней спячки. Названные суслики встречаются в западной части Северной Америки на высотах от 1500 до 3600 м над уровнем моря от севера Британской Колумбии до юга Калифорнии. Обычно их зимняя спячка длится три-четыре месяца, и за это время масса их тела значительно снижается. После спячки у них быстро растет потребление пищи и вес тела достигает отмечавшегося перед началом спячки в октябре. При изоляции в лабораторных условиях ритм зимней спячки и связанных с ней изменений в массе тела может сохраняться ряд лет даже при постоянных освещении и температуре (рис. 16.9). Если сыворотку крови от суслика в состоянии зимней спячки ввести неспящему животному, то последний также впадает в спячку (Pengelley, Asmundson, 1974). Этот факт служит веским доводом в пользу того, что основной физиологический механизм гибернации управляется эндогенными биологически-
| Рис. 16.9.Цирканнуальные ритмы изменения массы тела и периоды зимней спячки (закрашенные области) двух особей Citellus lateralis, содержавшихся в неизменных лабораторных условиях. (По Pengelley, Asmundson, 1974.) | 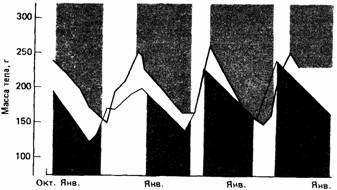
|
ми часами, захватываемыми ритмом внешних событий с помощью особого времязадателя. Это ясно показано на североамериканском лесном сурке, у которого также известен цирканнуальный ритм зимней спячки. Эти сурки были переправлены из восточной части США в Австралию - в новые для них условия освещения и температуры. Сурки за два года изменили нормальный ритм гибернации на обратный, приведя его в соответствие с местными условиями, несмотря на то, что в изобилии получали корм и воду и не страдали от понижения температуры.
Зимняя спячка похожа на акклиматизацию тем, что представляет собой длительное изменение физиологического состояния, позволяющее животному справляться с сезонными изменениями внешних условий. Цирканнуальные изменения физиологического состояния животного могут быть представлены в физиологическом пространстве. Годичные циклы размножения и зимней спячки являются в значительной степени врожденными, хотя фактически соответствуют общим сезонным изменениям в природе.
Миграция
Миграция из одного местообитания в другое на периодической или сезонной основе происходит у многих видов животных, в том числе у дневных бабочек, саранчи, лососей, птиц, летучих мышей и антилоп. Это не единственная форма миграции, поскольку у некоторых видов
миграция составляет часть обследования и колонизации новых территорий. Так, популяционный взрыв у норвежских леммингов (Lemmus lemmus) ведет к расселительной миграции, которая может охватить тысячи особей, обычно неполовозрелых самцов. Потоки леммингов устремляются вниз по склонам гор в долины. Многие тонут, пытаясь переплыть широкие водные пространства. Лемминги - хорошие пловцы и обычно не входят в воду, если не видят противоположного берега. Но порой давление множества скопившихся на берегу особей друг на друга таково, что некоторые из них вынуждены выплывать в открытое море.
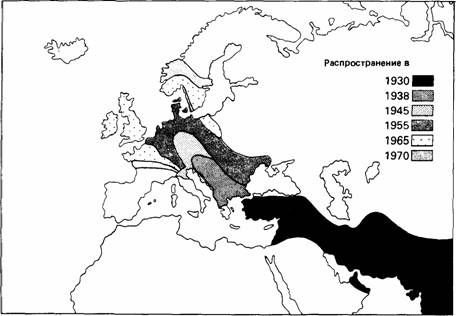
Заселение новых областей иногда достигается за счет постепенной миграции нескольких поколений, как, например, у кольчатой горлицы (Streptopelia decaocto) (рис. 16.10). Исследовательская миграция часто встречается у молодых позвоночных животных и при благоприятных условиях может обеспечить успешное расселение.
Периодические миграции возникают у многих видов в ответ на изменения внешних условий. Так, например, саранча Schistocerca gregaria заселяет в определенное время года безводные области. В зависимости от степени скопления эти насекомые развиваются в одну из трех имагинальных форм. Относящиеся к стадной форме особи собираются в плотные стаи, которые переселяются по ветру в области с низким барометрическим давлением, где скорее всего может идти дождь. Они летят днем и останавливаются, попав
|
| Рис. 16.10.Карта расширения ареала кольчатой горлицы (Streptopelia dccaocto) в XX столетии. (Из The Oxford Companion to Animal Behaviour. 1981.) |
|
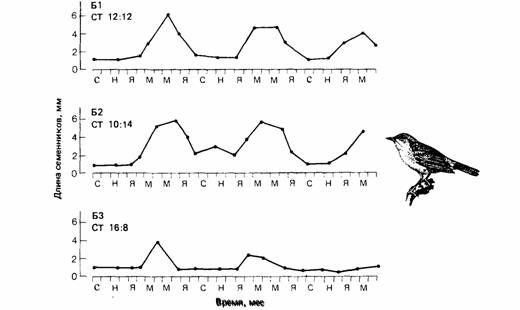
| Рис. 16.11. Цирканнуальные ритмы изменения длины семенников у садовых славок (Sylvia borin), содержавшихся в течение трех лет в неизменных фотопериодических условиях. СТ соотношение света и темноты. (По Berthold. 1974.) |
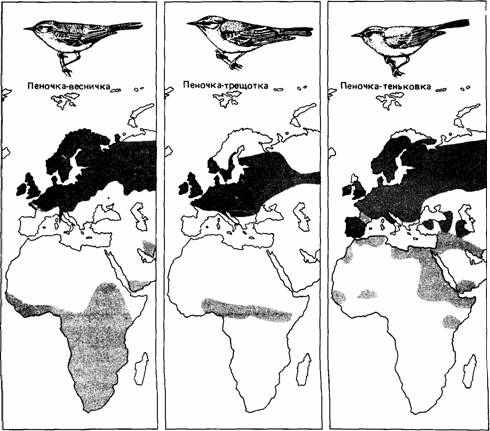
Рис. 16.12.Места гнездовий (темные участки), зимовок (бледные участки) и их перекрывания (черные участки) у некоторых мигрирующих пеночек. (По Schmidt-Koenig, 1979.)
во влажные условия. Здесь происходят половое созревание, спаривание и откладка яиц. Саранчевые стаи мигрируют по замкнутым сезонным маршрутам, но время генерации слишком коротко для того, чтобы отдельные особи могли завершить полный круг.
Некоторые сезонные миграции в отличие от периодических начинаются на основе цирканнуального ритма, а не в ответ на изменения внешней среды. У певчих птиц из родов Phylloscopus (Gwinner, 1971) и Sylvia (Berthold, 1973) наблюдали разное миграционное поведение. У таких типичных мигрантов на дальние расстояния, как садовая славка (S. borin), горная славка (S. cantillans) и пеночка-весничка (Ph. trochilus), происходят значительные сезонные изменения в массе тела, характере линьки, размере семенников, ночном беспокойстве и пищевых предпочтениях (рис. 16.11). Европейские популяции этих видов зимуют в Африке и мигрируют через Сахару (рис. 16.12). Если же они помещены в постоянные лабораторные условия через несколько дней после вылупления, то процессы, которые у свободноживущих птиц являются сезонными, сохраняют сезонность у особей тех же видов в неволе, хотя период колебаний этих процессов у них несколько короче календарного года.
Мигранты на средние расстояния, такие, как славка-черноголовка (S. atricapilla) ипеночка-теньковка (Ph. collybita), зимуют в Европе и Африке, причем масса тела и другие миграционные показатели у них меняются умеренно. В постоянных
лабораторных условиях у этих видов тоже происходят сезонные изменения. Белоусая славка (S. melanocephala) и провансальская славка (S. undata) являются частичными мигрантами, которые зимуют в средиземноморской области гнездования, a S. sarda balearica проводит весь год и эндемична на Балеарских и Питиузских островах в Средиземном море. У этих видов масса тела достаточно постоянна в течение всего года, а линька оперения на туловище продолжается долго и перемежается периодами ночного беспокойства. В лаборатории у них можно наблюдать некоторые проявления сезонных изменений, но с выраженными индивидуальными различиями (Berthold. 1974).
Вероятно, у видов этих двух родов происходят сезонные изменения миграционных показателей, интенсивность проявления которых коррелирует с миграционным поведением вида, а распределение во времени говорит о том, что начало миграции, возможно, частично регулируется эндогенным цирканнуальным ритмом. Подобные же сезонные ритмы установлены у некоторых видов птиц для таких физиологических процессов, как линька, отложение жира, миграционное беспокойство и размножение (Rutledge, 1974).
Получены также данные, что эндогенные факторы управляют не только началом миграционной активности, но и ее характером. У многих видов пути миграции характерны для определенных географических популяций. Так, например, белые аисты (Ciconia ciconia), гнездящиеся в Западной Европе, летят на места зимовок в Африку западным путем через Испанию и Гибралтар, а гнездящиеся в Восточной Европе избирают восточный путь, как показано на рис. 16.13. Неопытные молодые аисты, выращенные в неволе в Восточной Европе, но выпущенные на свободу в Западной, летят в юго-восточном направлении, характерном для птиц из Восточной Европы (Schuz, 1963). Такие же опыты с другими видами пернатых свидетельствуют о генетически обусловленном выборе направления миграции.
Чтобы достичь места зимовки, птица должна лететь не только в правильном направлении, но и на вполне определенное расстояние. Имеются данные (Gwinner, 1972) о расстояниях, которые преодолевают при первом перелете молодые славки, не сопровождаемые взрослыми птицами. Эти данные говорят о том, что у птиц существуют эндогенные часы, указывающие, сколько времени нужно лететь на каждом этапе перелета. Наблюдается хорошая корреляция между числом часов миграционного беспокойства птиц в неволе и расстоянием, нормально пролетаемым ими во время миграции (например, Berthold, 1973). Расстояние, эквивалентное одному часу такого беспокойства, может быть определено путем сравнения поведения птиц одной и той же популяции в неволе и во время перелета (Gwinner, 1972). Зная скорость полета мигрантов, можно вычислить расстояние, которое покрыли бы сидящие в клетке птицы, если бы летели в правильном направлении. Расчеты привели к результатам, близким к полученным при наблюдении над свободно мигрирующими птицами этой же популяции. Садовые славки (Sylvia borin), содержавшиеся в неволе во время первой осенней миграции, поворачивались на юго-запад в августе и сентябре и на юго-восток в октябре, ноябре и декабре (Gwinner. Wiltschko, 1978). Эти данные совпадают с «расписанием» естественной свободной миграции, показанным на рис. 16.14. Некоторые птицы перелетают много сотен миль над океаном, где ориентирами служат лишь несколько островков. Так, например, полярные крачки (Sterna paraclisea) размножаются в Арктике, а осенью мигрируют к антарктическим паковым льдам. Весной они совершают обратный перелет, хотя, вероятно, другим путем. Известно, что некоторые особи пролетают 9000 миль (14500 км). Подобным же образом тонкоклювый буревестник (Puffinus tenuirostris) размножается в юго-восточной Австралии и мигрирует на Аляску через Японию. Обратное путешествие совершается вдоль западного берега Северной Америки (рис. 16.15).
Дата добавления: 2016-06-18; просмотров: 2443;
Поиск по сайту
Узнать еще
- II. КРОВЬ КАК ВНУТРЕННЯЯ СРЕДА ОРГАНИЗМА. ФИЗИОЛОГИЯ ЭРИТРОЦИТОВ И ЛЕЙКОЦИТОВ.
- II. Поведение веществ в магнитном поле.
- АГРЕССИВНОЕ ПОВЕДЕНИЕ
- Агрессивное поведение. Основные положения
- АДДИКТИВНОЕ ПОВЕДЕНИЕ НЕСОВЕРШЕННОЛЕТНИХ
- Анатомия и физиология
- Анатомия и физиология в сочетании с толкованием аномалий в общем, не симптоматическом ключе, являются основами остеопатии.
- Анатомия и физиология глотки. Острый и хронический фарингиты.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
