Генетический материал
Мендель не пользовался термином ген. Он писал о видимых (фенотипических) признаках живых организмов и постулировал существование элементов, или факторов, наследственности, которые ведут себя как дискретные единицы, или корпускулы. Когда в 1900 г. работы Менделя привлекли внимание научного мира, ученые уже предполагали, что наследственный материал содержится в хромосомах. Интенсивные исследования подтвердили всеобщность закона Менделя, хотя, как отмечалось ранее, были выявлены и исключения. В 1905 г. Бэтсон дал этой области исследований название «генетика», а термином «ген» датский ботаник Йоганнсен (Johannsen) в 1909 г. назвал менделевские элементы наследственности. Йоганнсен установил также и важное различие между генотипом и фенотипом.
Морган называл генами самые маленькие единицы рекомбинации, расположенные на хромосоме, как бусы на нитке. Сцепленные гены могут рекомбинировать путем кроссинговера. Ген рассматривают также и как функциональную единицу, и как единицу мутации. Таким образом, генная мутация представляет собой мельчайшее изменение генотипа, благодаря которому меняется фенотип. В начале нашего столетия концепция гена была единой. Однако успехами в понимании химической природы гена это единство было нарушено. В работах Эвери (Avery, 1944), Уотсона и Крика (Watson, Crick, 1953) было показано, что генетический код записан на молекулах ДНК (дезоксирибо-нуклеиновой кислоты).
|
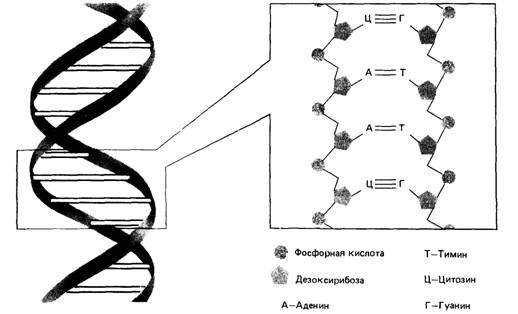
| Рис. 2.8. Схематическое изображение двойной спирали ДНК. Каждая цепь образована чередующимися остатками молекул сахара (дезоксирибозы) и фосфатной группы. Эти две цепи соединяются слабыми связями между нуклеотидными основаниями: аденином (А), тимином (Т), цитозином (Ц) и гуанином (Г). |
Молекула ДНК состоит из двух цепей (рис. 2.8), каждая из которых в свою очередь состоит из отдельных нуклеотидов. Эти нуклеотиды являются элементами генетического кода. ДНК выполняет две функции. Она воспроизводит себя, обеспечивая наследственную связь между поколениями, и предоставляет информацию, необходимую для создания тысяч различных белков. Точно так же как ДНК образована из последовательности нуклеотидов, белок образован из небольших молекул аминокислот. Каждая возможная комбинация трех нуклеотидов (кодон) из четырех нуклеотидных типов определяет один из 20 типов аминокислот (существуют также кодоны, которые детерминируют обрыв синтеза белковой цепи или несут другую информацию). Сама цепь ДНК не покидает ядра, поэтому сначала она транскрибируется в матричную, или информационную, РНК (рибонуклеиновую кислоту), которая перемещается из ядра в цитоплазму клетки, где находится запас аминокислот, из которых синтезируются функциональные белки. Ген в данном случае просто участок ДНК, который включает всю последовательность нуклеотидов (или РНК), кодирующих один белок. В хромосомах любого сложно организованного животного помимо десятков тысяч генов имеются также и длинные участки ДНК, которые не кодируют синтез каких-либо известных генных продуктов. Генные мутации могут вызывать различные типы нарушения в молекуле ДНК. В частности, могут происходить утраты (делеции) или замены нуклеотидов. Если такие изменения происходят в начале гена, то наблюдается неправильное считывание триплетов вдоль всей цепи в результате чего образуется совершенно другой нефункциональный белок. Если вставка или утрата участка произошла в конце гена, то начало белковой цепи строится правильно, а конец ее изменен тем или иным образом. Если изменения не коснулись активных участков молекулы, то некоторые ее функции могут сохраниться. Другой тип мутаций - замена оснований, т. е. замена одного нуклеотида другим, в
результате чего образуется новый триплет, кодирующий другой белок, или триплет, который разорвет активные части цепи. Мутации возможны и в генах-регуляторах, контролирующих активность других генов.
Хромосомные мутации включают рекомбинацию генов, так что они располагаются на хромосоме в другом порядке или попадают в другие хромосомы. Этот тип мутаций не изменяет конкретного гена, но может влиять на способ образования белков, поскольку порядок расположения генов в хромосоме - важный аспект экспрессии генов.
Частота мутаций в отдельно взятом гене довольно низка. Однако число генов у особи очень велико, а число генов в генофонде популяции поистине грандиозно. Поэтому в целом по популяции мутации происходят часто, однако у какой особи возникнет мутация - определяется чистой случайностью. В подавляющем большинстве мутации вредны и вызывают гибель особи на ранних стадиях развития. Лишь некоторые мутации позволяют потомству выжить. Такие мутации увеличивают генотипическую изменчивость, являющуюся объектом действия естественного отбора, но они не определяют направление эволюции.
ДЛЯ ЗАПОМИНАНИЯ
1. Действие естественного отбора зависит от наличия генотипи-ческой изменчивости в популяции. Те особи, которые лучше всего приспособлены к окружающей среде, имеют возможность оставить более многочисленное потомство. Изменчивость возникает в процессе мейоза, а также в результате хромосомных и генных мутаций.
2. В процессе мейоза происходит перераспределение генетического материала в соответствии с законами Менделя. Это может привести к появлению новых генных комбинаций в ряду поколений.
3. Хромосомные мутации также происходят во время мейоза, но они происходят не систематически. Это ошибки мейоза, которые случайны и дают новые генетические комбинации.
4. Генные мутации-это изменения в химической структуре гена. Они происходят редко и в основном вредны для особи. Однако в результате некоторых мутаций появляются полезные новшества, обеспечивающие особи эволюционные преимущества.
Рекомендуемая литература
Ayala F.J., Kiger J.A. (1984). Modern Genetics, 2nd edition, Benijamin/Cummings, Menlo Park, California.
Развитие поведения
Типы поведения животных складываются в результате сложного взаимодействия внешних стимулов и состояния внутренней среды организма. Однако любой тип поведения определяется тем, каким образом животное обрабатывает поступающую к нему информацию. Внутренние системы обработки информации формируются в процессе развития от оплодотворенного яйца к зародышу и взрослому животному; этот процесс называется онтогенезом. Изучая онтогенез, мы можем выявить способы взаимодействия генетической информации и информации, полученной из окружающей среды (или путем научения), что в конечном счете формирует соответствующий тип поведения животных.
Онтогенез
Почти все живые организмы построены из одних и тех же веществ, причем различия между видами определяются различиями в способах объединения этих основных веществ. Механизмы, контролирующие такое объединение, изучены слабо, однако известно, что гены контролируют развитие, детерминируя синтез белков, которые регулируют сложную организацию процессов онтогенеза.
Однако эти регуляторные белки эффективны лишь настолько, насколько развивающийся организм чувствителен к информации, которую они несут. Действие регуляторной молекулы основано не на создании некой системы, соответствующей какому-то определенному плану, напротив, эта молекула вызывает специфическую реакцию в клетках, на поверхности которых находятся рецепторы для регуляторных молекул. Таким образом, ответ развивающегося зародыша на продукт регуляторного гена зависит от внутренней организации зародыша ко времени образования этого регулятора. Кроме того, этот регулятор функционирует только при определенных условиях внутренней среды организма. В качестве примера рассмотрим сначала некоторые аспекты эмбриогенеза у видов с относительно простым поведенческим репертуаром.
Aplysia - лишенная раковины морская улитка, масса которой может достигать 4,5 кг. Как типичная улитка, Aplysia - настоящийгермафродит. Оплодотворение у нее внутреннее, и обычно образуются цепи, включающие до 10 и более копулирующих животных, причем каждая улитка оплодотворяет соседнюю, в то время как сама оплодотворяется другой. После оплодотворения Aplysia откладывает более миллиона яиц, соединенных в длинный шнур. Сначала шнур выталкивается из репродуктивного мешка сокращением мышц последнего. Когда шнур достигает достаточной длины, улитка захватывает его конец в складку верхней губы и, двигая головой взад и вперед, помогает высвобождению яиц из репродуктивного мешка (рис. 3.1). Специальная железа секретирует клейкое вещество, с помощью которого шнур склеивается, образуя более компактную массу в виде беспорядочного мотка. Наконец, улитка крепко прижимает головой эту массу яиц к твердому субстрату (рис. 3.1), где они и остаются до вылупления. Вся эта серия последовательных действий при откладке яиц-жест-ко зафиксированный поведенческий стереотип. Во время этого процесса у животного увеличивается частота сокращений

|
| Рис. 3.1. Поведение Aplisvu, откладывающей яйца. Лента яиц в защитной оболочке выталкивается из полового протока, выходное отверстие которого расположено на боку тела. Животное захватывает этот шнур в складку верхней губы (слева) и прикрепляет его к субстрату (справа). (По Scheller, Axel, 1981.) |
сердца и частота дыхательных движений, оно полностью прекращает питаться и передвигаться.
Шеллер и Аксель (Scheller, Axel, 1981) обнаружили, что эта последовательность действий контролируется генами на разных уровнях, как во время эмбриогенеза, так и в процессе ее осуществления. Клетки, которые дают начало всей нервной системе аплизии, являются потомками нескольких клеток, располагающихся в стенке тела зародыша. На ранних стадиях развития эти преднервные клетки синтезируют специфический белок, названный гормоном откладывания яиц (ELH-egg-laying hormone). Полагают, что все эти преднервные клетки являются потомками единственной клетки зародыша, в которой реализуется способность соответствующего гена кодировать синтез ELH. Во всех других клетках зародыша также имеется этот ген, но в них он не функционирует. Клетки, синтезирующие ELH, продолжают делиться и перемещаются к своему окончательному местоположению в развивающемся организме, где дифференцируются в нервные клетки взрослой особи. Поэтому у взрослой улитки некоторые нервные клетки наследуют способность синтезировать ELH, а другие - нет.
Ко времени полового созревания все эти системы улитки занимают соответствующее им местоположение. В надлежащее время начинают вырабатываться другие гормоны, которые и приводят клетки, синтезирующие ELH, в состояние полной работоспособности. Выделяющийся ELH заставляет возбуждаться соседние нейроны (т. е. в данном случае гормон действует как нейромедиатор). Этот гормон циркулирует и в крови, заставляя сокращаться специфические мышечные волокна и действуя, таким образом, как гормон; в результате приводится в действие сложный комплекс координированного поведения откладки яиц. На примере таких фиксированных действий отчетливо видна вся важность координации процессов эмбриогенеза, активности генов, действия нейромедиаторов, нервов и мышц, приводящей к реализации поведения, столь существенного для размножения улитки.
У аплизии гены непосредственно контролируют развитие стереотипных форм поведения. И если гены на каком-то уровне (например, в эмбриогенезе, когда происходит закладка общего плана строения) контролируют становление поведения в целом, то различные средовые факторы могут в разной степени модифицировать те или иные формы поведения, появившиеся в эмбриогенезе. Фенотипический эффект одних и тех же генов может быть различен в зависимости от того, в каких условиях развивается животное. Например, у ракообразного Gammarus глаза обычно красного цвета. Форд и Хаксли (Ford, Huxley, 1927) обнаружили у Gammarus мутацию, которая влияет на скорость образования глазного пигмента во время определенной стадии онтогенеза. Если на этой стадии развития температуру
окружающей среды повысить до некоторого уровня, то глаза остаются красными, при более высоких температурах они становятся шоколадно-коричневыми, а при промежуточных температурах - промежуточного цвета. Учитывая огромную сложность биохимических процессов, принимающих участие в экспрессии генов, в процессах роста и дифференцировки клеток (Ham, Veomett, 1980), не приходится удивляться, что среда, в которой протекают данные процессы, влияет на развитие этих клеток. Присущие животным стабилизирующие, или регуляторные механизмы в какой-то степени исправляют отклонения и контролируют скорость и направление процессов развития. Иногда одна и та же структура или один и тот же тип поведения развиваются разными путями - явление, известное под названием эквифинальностъ. Тем не менее отклонения от обычного пути развития неизбежны. В некоторых случаях эти отклонения могут быть летальными, в других же они приводят к возникновению различий между особями, несущими один и тот же генотип.
Некоторые ранние этологи считали, что гены непосредственно детерминируют поведение. Так, Лоренц (Lorenz, 1965) полагал, что генетическое детерминирование аналогично архитектурному проекту здания. Генетический проект представляет собой некий план создания взрослого животного. Признавая, что для строительства здания нужны кирпичи, раствор и рабочая сила, Лоренц определил четкие различия между условиями, необходимыми для воплощения проекта в здание, и информацией, которая содержится в проекте и от которой зависят характеристики готового сооружения. Исследователи, критиковавшие эту точку зрения (например, Lehrman, 1970), считали, что если архитектурный проект изоморфен строению, которое он представляет, то этого нельзя сказать в случае генетического проекта. Ведь между измерениями, указанными на архитектурном проекте, и измерениями построенного здания существует однозначное соответствие, какого не может быть в онтогенезе. Даже если верно то, что в каждом гене закодирован определенный ключевой фермент, все же это сильно отличается от проекта. Представление о том, что информацию, получаемую в процессе развития от генов, можно отделить от информации, получаемой из окружающей среды, было критически воспринято Хеббом (Hebb, 1953), который считал, что спрашивать, насколько данный тип поведения зависит от генетических, а насколько от средовых факторов, - это все равно, что спрашивать, насколько площадь поля зависит от его длины, а насколько - от ширины.
Большой интерес исследователей к изучению онтогенеза поведения объяснялся их желанием продемонстрировать разнообразие путей, по которым развиваются различные типы поведения. Оказалось, что существует целый спектр процессов, различающихся по влиянию на них факторов окружающей среды, - от относительно независимых до полностью зависимых.
Генетический детерминизм ранних этологов привел к признанию того, что влияние генов и влияние окружающей среды сложным образом связано друг с другом, однако связь эта не такая, как полагал Лоренц (Lorenz, 1965), - связь проекта и построенного на его основе здания; скорее она носит эпигенетический характер, т.е. каждая стадия развития создает основу для следующей стадии, а не предопределяет ее. По мнению Брауна (Brown, 1975), процессы эпигенеза суммарно можно представить следующим образом. Начиная с оплодотворенного яйца, или зиготы (P1), фенотип организма во время следующей стадии развития (Р2) будет определяться генами, которые детерминируют его рост и дифференцировку в данный период (G1), и условиями среды (E1), в которой протекает развитие; т. е. зигота плюс гены плюс окружающая среда будут определять фенотип на следующей стадии:
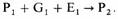
Фенотип (Р3) на следующей стадии развития будет определяться взаимодействием фенотипа Р2, генов G2 и условий окружающей среды (Е2); это приведет к развитию фенотипа Р3:
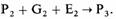
Эта запись показывает, что все три элемента - ген, окружающая среда и исходный фенотип - играют определенную роль в нормальном развитии. Развитие может достичь следующей стадии только в том случае, если развивающееся животное обладает соответствующим фенотипом, если у него синтезируются правильные генные продукты и если условия среды не выходят за рамки определенных границ.
На ранних стадиях развития (P1 + G1 + Е1 --> Р2) средовой компонент E1 представляет собой в основном биохимическое окружение, в котором находится ранний зародыш. На более поздней стадии развития (Р2 + G2 + Е2 --> Р3) средовой компонент Е2 может быть представлен веществами яйца. После вылупления из яйца или рождения окружающая среда животного включает информацию, получаемую его органами чувств, а также биохимические вещества, получаемые в процессе питания и другими способами. В этой ситуации может иметь место научение, которое постоянно влияет на поведенческий фенотип (Р3) животного.
Связь между врожденным и приобретенным поведением удобно изучать, сравнивая птенцов выводковых (у которых вылупление происходит на поздних стадиях развития) и птенцовых (у которых вылупление имеет место на ранних стадиях) видов птиц. Если сравнивать птенцов этих птиц на одной и той же стадии развития, например на стадии, когда появляются первые перья, то оказывается, что птенец выводковой птицы находится в это время еще в яйце, тогда как птенец птенцовой птицы уже вылупился. Ко времени вылупления выводковой птицы молодая птенцовая имела возможность уже многому научиться, тогда как выводковый птенец учиться еще и не начинал. И что особенно интересно - мы обнаружили, что выводковая птица может обладать такими врожденными навыками которые птенцовой птице приходится осваивать. Например, белоголовая воробьиная овсянка, относящаяся к птенцовым птицам, заучивает видоспецифическую песню с первых дней своей жизни в гнезде. Напротив, птицы, относящиеся к выводковым, проходят ранние стадии развития в яйце и при вылуплении уже обладают способностью к нормальной вокализации, хотя раньше никогда не слышали звуки, издаваемые родителями.
Мы склонны думать, что происходящие внутри яйца события представляют собой лишь физические процессы созревания; иными словами, мы считаем, что факторы, влияющие на этот процесс, предопределены. Однако мы не должны забывать, что нормальная среда белоголовой воробьиной овсянки также предопределена в том смысле, что в критический для научения песне период птенцы в гнезде обычно слышат песню своего вида.
На последующих стадиях развития Р3 + G3 + Е3 --> Р4. К этой стадии влияние приобретенного опыта будет заметно как у птенцовых, так и у выводковых птиц. Поведение, которое несомненно складывается под влиянием опыта, нельзя назвать «врожденным». Однако здесь есть небольшое принципиальное отличие от ситуации, складывающейся на ранних стадиях развития. Степень предопределенности средовых факторов зависит в основном от изменчивости окружающей среды. Внутри яйца среда обычно изменяется незначительно, поэтому мы склонны считать поведение только что вылупившегося птенца врожденным. Однако вне яйца среда также может изменяться очень незначительно, и это зависит от экологических условий, типичных для вида. Таким образом, постнатальное поведение птенцовых птиц может быть точно так же предопределено, как и постнатальное поведение выводковых. Если с эволюционной точки зрения важно, чтобы развитие давало какой-то конкретный конечный результат, как это отчетливо видно в случае с песней у воробьиных птиц, то мы можем ожидать, что влияние окружающей среды будет таким, чтобы обеспечить этот результат.
Дата добавления: 2016-06-18; просмотров: 2199;
Поиск по сайту
Узнать еще
- Audit Trail - Материалы для проведения аудиторской проверки
- III. Проводниковые материалы
- III. ТИТАН – МАТЕРИАЛ ДЛЯ СОВРЕМЕННОЙ СТОМАТОЛОГИИ
- III.1. КИНЕМАТИКА МАТЕРИАЛЬНОЙ ТОЧКИ
- VII. Педагогические технологии на основе дидактического усовершенствования и реконструирования материала
- Yandex_direct_print() Материалы кровельных покрытий
- Аварийное снабжение и материалы
- Автомобильные топлива, смазочные материалы и специальные жидкости.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
