И ЭКВАТОРИАЛЬНЫЕ ЛЕСА
Географическое распространение,
гигротермический и геохимический режимы
Вечнозеленые влажные тропические и экваториальные леса распространены вблизи экватора в виде прерывистой полосы. Основные массивы этих лесов (часто называемых также гилеей, сельвой, дождевым тропическим лесом, влажнотропическим лесом) на равнинах не выходят к северу и югу далее 10-й параллели и сосредоточены в областях настоящего экваториального климата. Наиболее крупный участок в Южной Америке приурочен к бассейну Амазонки, в Африке - к бассейну Конго и южному побережью Гвинейского залива, в Азии - к Малакке, Суматре, Калимантану (Борнео), Филиппинам. Большая часть Новой Гвинеи также покрыта дождевым тропическим лесом, включая его горные варианты. Сравнительно небольшими языками или даже островами вечнозеленые леса проникают до тропика Рака в странах Карибского моря, а в Азии - на востоке Индии, в Бангладеш, Непале, Бирме и на крайнем юго-западе Китая - даже переходят его. Вдоль западного побережья Индии и в Шри-Ланке, а также южнее экватора, в Квинсленде (по восточному побережью Австралии), на ряде островов Океании, в Бразилии и на Мадагаскаре также растут вечнозеленые тропические леса (включая и самые влажные варианты муссонных лесов), достигающие местами тропика Козерога, а южнее переходящие уже в субтропические варианты.
Внутри очерченного нами ареала первичная гилея сохранилась далеко не повсеместно. Большие пространства заняты вторичными лесами или даже заменены злаковыми сообществами. Такого рода деградация отмечается в прибрежных «мату атлантику» Бразилии, во многих районах Центральной Америки, Мадагаскара, Западной Африки, Индии, Индокитая. Относительно лучше пока еще сохранились приэкваториальные леса востока Амазонки, Конго и в отдельных областях Юго-Восточной Азии.
Наиболее характерная черта гигротермического режима вечнозеленых тропических и экваториальных лесов - круглогодичное достаточное увлажнение и ровные высокие температуры. Среднегодовая температура обычно близка к +25...+26°С, при этом различия между среднемесячными величинами редко превышают 5° С, а максимальные температуры доходят лишь до +33...+35°С. Вместе с тем суточные амплитуды нередко равны 10°С (они колеблются в пределах 3-16°С в разных районах). По мере удаления от экватора среднегодовые температуры могут (хотя и не всегда) снижаться до +20°С, это же происходит и при подъеме в горы. Общая годовая сумма осадков колеблется очень сильно, хотя всегда превышает 2000 мм. Главным фактором выступает не столько общая сумма осадков, сколько ритм их выпадения - в особенности важны продолжительность сухих периодов и их резкость. В течение большей части года количество осадков превышает испарение. Обратное соотношение наблюдается не дольше 1-2 месяцев в году.
Для экологических и биогеографических целей весьма подходящим надо признать метод характеристики климатических условий с помощью климадиаграмм. Основа такой климадиаграммы формируется из кривой среднемесячных температур и среднемесячных осадков, при этом одно деление вертикальной оси соответствует 5°С и 10 мм. Пространство, ограниченное снизу температурной кривой, а сверху - кривой осадков, соответствует периоду с достаточным увлажнением; обратное расположение кривых выявляет засушливую часть года. Периоды со среднемесячным количеством осадков выше 100 мм изображают в более мелком (в 10 раз) масштабе и заливают черным цветом (это сезон избыточного увлажнения). На диаграмме указывают также периоды с регулярными и эпизодическими морозами (если они есть), среднегодовые температуры (включая максимумы и минимумы) и осадки, абсолютные максимумы и минимумы температур, температурную среднесуточную амплитуду и, наконец, абсолютную высоту станции и ее название. На рисунке 11 показаны климадиаграммы некоторых пунктов, лежащих в разных областях распространения тропических дождевых лесов.
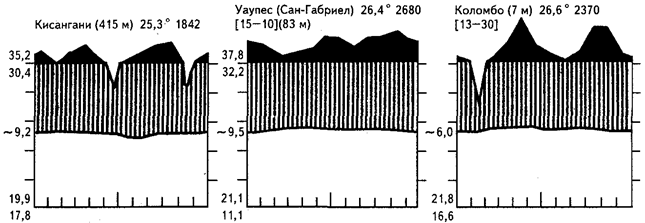
Рис. 11. Климадиаграммы некоторых районов распространения
тропических дождевых лесов на равнинах
К сожалению, на климадиаграммах для влажных тропиков слабо выделяется один из важнейших показателей присущего им климата - характер (часто очень регулярный) суточных колебаний различных метеорологических факторов, которые обычно превосходят амплитуду среднемесячных значений. Так, например, в Богоре на Яве (здесь расположен знаменитый влажнотропический ботанический сад) средние месячные температуры колеблются лишь в пределах +24,3...+25,3°С. Однако в солнечные ноябрьские дни в течение суток отмечены изменения в пределах +23,4...+32,4°С. Даже в самые влажные месяцы амплитуда колебаний в течение суток достигает 6-7 "С. Обычно до полудня светит солнце, а во второй половине дня бывают ливни.
Отмеченные выше особенности влаго- и теплообеспеченности под пологом леса значительно смягчаются, колебания влажности и температур здесь еще более плавны, но в верхней части крон они увеличиваются. Еще заметнее сверху вниз изменяется освещенность — на поверхности почвы она составляет всего 0,1%-1% (чаще 0,5%) от полного дневного света. Как и в широколиственных лесах умеренных широт, уровень освещенности в 0,5% уже близок к пределу для произрастания теневыносливых цветковых растений.
В пределах даже одинаковых условий увлажнения и температур почвенно-геохимические условия могут существенно меняться, главным образом это связано с приуроченностью к тем или иным элементам рельефа и к различным материнским породам. Тем не менее большая часть почв дождевых тропических лесов обладает рядом общих специфических свойств, важных для жизнедеятельности в первую очередь растительных организмов. Так, преобладают почвы глинистого или суглинистого состава, обедненные основаниями и элементами минерального питания. Реакция почвенных растворов всегда кислая, окраска почв часто ярко-красная или желтоватая, что связано с избытком окислов железа.
Многие почвы влажных тропиков имеют весьма древний возраст. Это обстоятельство, а также высокие температуры и влажность приводят к формированию очень мощной коры выветривания, которая может достигать толщины 20-30 м. Благодаря выносу ряда веществ, в первую очередь оснований и кремнезема, происходит относительное накопление полуторных окислов алюминия и железа. Несмотря на мощную толщу коры выветривания, сама почва (т. е. населенная организмами верхняя часть коры выветривания) имеет мощность всего до 2 м. В большинстве случаев корни растений в гилее вообще не проникают глубже 1 м (это связано отчасти с плохой аэрацией). Из-за интенсивно идущих процессов разложения накоплений больших запасов подстилки, а также существенных накоплений гумуса в верхних почвенных горизонтах не происходит. Органическое вещество часто находится в почве в растворимом виде, общее количество его обычно составляет 2-5%. Почвенные значения рН колеблются в пределах 4,5-5,5, при этом в самых верхних горизонтах и в скоплениях опада среда еще более кислая (до 3,5). Карбонаты кальция и магния практически отсутствуют, в многочисленных ручьях и речках вода мягкая, почти обессоленная, имеющая желтоватый или янтарный оттенок из-за растворов дубильных и гуминовых веществ. Вода почти лишена микроорганизмов. Элементы минерального питания весьма скудно представлены в большинстве почв, исключая молодые почвы на вулканических отложениях.
Крайняя бедность почв в сочетании с интенсивным промывным режимом сквозь мощную кору выветривания кажется несовместимой с богатством и пышностью органического мира влажных тропиков. Этот парадокс разрешается просто, если обратить внимание на особенности круговорота веществ в экосистеме. Дело в том, что большая часть биогенных веществ (прежде всего элементов минерального питания фотоавтотрофов) сосредоточена в самих живых организмах. При их отмирании происходит быстрое разложение и элементы минерального питания моментально усваиваются растениями и таким образом снова включаются в живую биомассу. Раздумывая об этой удивительной особенности влажных тропических лесов, Г. Вальтер задает вопрос: «Как же первоначально образовался запас питательных веществ, сосредоточенный в фитомассе?» Возможен, по Г. Вальтеру, следующий ответ: «Исходный запас питательных веществ образовался в то время, когда мощность коры выветривания была не столь великой и корни растений непосредственно соприкасались с коренной породой. В тропиках этот период первоначального накопления следует искать в весьма далеком геологическом прошлом»1. В свете указанных фактов не удивительно, что после сведения первичных тропических лесов во многих районах вторичные леса даже через многие десятилетия не достигают пышности и высоты первичного дождевого леса. При сельскохозяйственном использовании таких участков урожаи на них падают почти до нуля уже через несколько лет и для посевов необходимо бывает расчистить и выжечь новый участок.
Впрочем, вторичный лес способен через некоторое время в значительной степени восстановить плодородие почвы за счет того, что в его составе главная роль принадлежит бобовым, способствующим накоплению азота в почве благодаря деятельности азотфиксирующих клубеньковых бактерий.
На песчаных почвах и на кислых коренных породах ферралитовые почвы не образуются, а протекают процессы подзолообразования. В почвах при этом выделяются элювиальные и иллювиальные горизонты, часто присутствует грубогумусовый подстилочный горизонт. В такого рода эдафических условиях всегда встречаются особые типы растительности, например монодоминантные леса, кустарниковые заросли и даже саванны. Меньшее богатство растительности на таких почвах можно связать с отсутствием достаточного количества питательных веществ даже при начальных фазах выветривания коренных пород. В районах с большим распространением выщелоченных песков вода в реках имеет характерный коричневый цвет. В частности, именно этим объясняется название притока Амазонки Риу-Негру, вода в котором окрашена коллоидами «сырого» гумуса в темный цвет. Такой же цвет имеют и реки, вытекающие из тропических торфяных болот, поросших лесом. В Бразилии подобные водотоки нередко называют «агуа прэта» - вода-негритянка, в отличие от мутной и бесцветной «агуа бранка» - белая вода.
Сообщества фотоавтотрофов
Сообщества фотоавтотрофов (зеленых растений) формируют первый трофический уровень, продукция которого одновременно представляет собой и общую биологическую продукцию экосистемы. Описание этой группы организмов практически означает характеристику растительности, ибо доля паразитических и сапротрофных групп весьма мала в сравнении с долей зеленых растений. Сообщества фотоавтотрофов из-за своего подавляющего доминирования по массе определяют общий облик, физиономию всего биотического сообщества, его пространственное построение, его средообразующую роль.
Нередко вечнозеленые тропические леса изображаются в описаниях очевидцев как непролазные «джунгли», в которых шага нельзя ступить без прорубания сквозь стену лиан и ветвей. Повсюду встречаются перистые пальмы, травы с гигантскими листьями, папоротники, бамбуки, густая высокая трава, колючие кусты... Однако этот экзотический для жителя умеренных широт пейзаж всего лишь вторичная растительность, буйно развивающаяся после сведения первичных девственных лесов. Селения и пашни прячутся в этом буйстве зелени, и даже весьма населенные районы могут поэтому показаться мало освоенными. В действительности густые заслоны из лиан, трав, пальм и бамбука растут в основном лишь по опушкам первичного дождевого леса, по берегам рек, на крутых склонах обрывов. Сам же первичный лес вполне проходим. Под его пологом в первую очередь видна колоннада высоких стройных стволов деревьев с отдельными обвивающими их лианами; кроны смыкаются уже над головой; поверхность почвы местами оголена и может быть глинистой и скользкой; обращают на себя внимание протянувшиеся по поверхности корня, многие из них дисковидные, сжатые с боков. Снизу трудно разобрать детали строения ветвей и листьев, вообще плохо просматриваются верхние ярусы, более всего насыщенные зеленью. Несмотря на богатство видами, внешний облик дождевого тропического леса у наблюдателя, не обладающего специальными знаниями, может оставить впечатление однообразия, так как в нем господствуют очень похожие друг на друга деревья с темно-зелеными листьями, напоминающими листья лимона или лавра. Однако вряд ли найдется ботаник, обманувшийся этим впечатлением.
Во влажных тропиках существует множество вариантов, уклоняющихся от весьма абстрактно «обобщенного типа» первичного дождевого леса, описанного в книгах. В то же время большинство структурных особенностей этих лесов в точности повторяется в разных районах, на разных континентах.
Наиболее типичные равнинные дождевые леса в среднем несколько выше лесов умеренных широт. Самых крупных деревьев, достигающих в высоту 60 м, обычно немного, всего несколько на сотни гектаров. Эти деревья возвышаются над весьма разреженным и неровным верхним ярусом, не превышающим, как правило, 50 м (в среднем около 35 м). Ниже пространство заполнено кронами разнообразных во высоте деревьев. В результате выделение древесных ярусов очень затруднено. Некоторые исследователи выделяют множество древесных ярусов, другие же говорят об отсутствии ярусности и равномерном заполнении всего пространства кронами. Непосредственное наблюдение ярусности в ненарушенном дождевом лесу обычно невозможно. Полагаться на картину, открывающуюся по опушкам и берегам водоемов, нельзя. Наиболее объективным оказывается метод профильной диаграммы, при котором на основании точных промеров рисуют в масштабе вертикальный разрез леса длиной не менее 60 м при ширине около 8 м. Приведенная серия таких профильных диаграмм для разных регионов и разных по составу дождевых лесов доказывает весьма большое разнообразие ярусной структуры (рис. 12, 13).
Лучше всего обычно выделяется ярус невысоких деревьев, в некоторых типах - и другие ярусы древостоя, часто же даже на вертикальных проекциях уловить границы между ярусами практически невозможно. В наиболее типичном смешанном дождевом лесу (состоящем из многих видов без выраженного доминирования 1-2 видов) выделяются 3 древесных яруса: ярус А - самый верхний, В - средний, С - нижний древесный. Обычно ярусы А та. В весьма постепенно переходят друг в друга, но даже и в этом случае возможно их вычленение и определение средней высоты. Не всегда это легко сделать на глаз, но можно воспользоваться линейкой и измерять по диаграмме через небольшие интервалы высот суммарные величины разрывов между кронами. Полученные цифры легко проанализировать или нанести их на график (рис. 14).
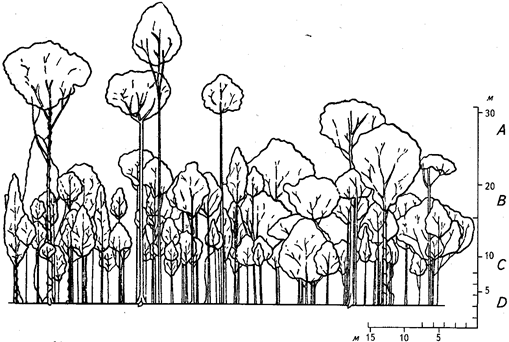
Рис. 12. Профильная диаграмма смешанного вечнозеленого тропического леса
с преобладанием видов семейства диптерокарповых. Калимантан, гора Дьюлит
(по П. Ричардсу, 1961)
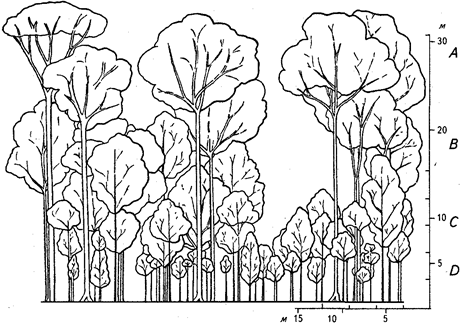
Рис. 13. Профильная диаграмма смешанного вечнозеленого леса Морабалли-Крик, Гайана. Показаны деревья высотой более 4,6 м (по П. Ричардсу, 1961)

Рис. 14. Общая высота деревьев и высота расположения самых нижних листьев кроны на участке близ Морабалли-Крик, Гайана. Сплошная линия — распределение деревьев по высотам, прерывистая — высота нижних пределов крон (по П. Ричардсу, 1961)
Выделение указанных трех древесных ярусов во всех случаях весьма целесообразно, так как дает возможность рационально организовать анализ и описание первичных материалов для более общих суждений о строении высокоствольных лесных сообществ.
Нередко при описаниях ярусности тропических лесов апеллируют к данным по распределению высот деревьев. Делать это можно лишь с большими оговорками, ибо ярусы древостоя формируются прежде всего не за счет числа стволов того или иного высотного класса, а за счет формы и величины крон разных по высоте деревьев. Весьма малочисленные деревья, достигающие большой высоты, чаще всего имеют развитую в ширину крону и поэтому могут сформировать хорошо выраженный ярус. Все же объективные данные о распределении деревьев по высотам показывают обычно 1-2 явно преобладающих класса высот, причем чаще наиболее резко выражен ярус С, который и на большинстве профильных диаграмм виден весьма четко.
Выше сомкнутого яруса С (высота его обычно близка к 8-10 м) деревья ярусов А и Б обычно не образуют сплошного полога, а встречаются куртинами разного размера. Ниже этого яруса выделяют ярус -D, представленный главным образом подростом древесных пород и высокорослыми травами. Еще ниже расположен напочвенный ярус Ё из трав и сеянцев деревьев. Независимо от принадлежности и тому или иному виду деревьев в ярусе А преобладают широкие, почти зонтиковидные кроны. В ярусе С господствуют узкие, пирамидальные, суживающиеся кверху кроны. В ярусе В кроны промежуточны, чаще всего они слегка вытянуты снизу вверх и имеют овальную форму. В монодоминантных формациях дождевых лесов очень часто ярус В практически выпадает, благодаря чему остаются очень четко выраженные ярусы А и С. На протяжении своей жизни высокие деревья поочередно входят в ярусы С, В, А, и, соответственно, изменяется форма их кроны.
По сравнению с лесами умеренных широт деревья в дождевом тропическом лесу имеют более тонкие, вытянутые, устремленные вверх стволы. Особенно это выражено у представителей яруса А, что также подчеркивается отсутствием ветвей в нижней части. Значительное отличие облика влажнотропических деревьев от наших определяется и тем, что первые имеют меньшую степень разветвленности. Если для европейских лиственных деревьев ветвление до пятой - восьмой степени весьма обычно, то в гилее чаще всего встречается лишь вторая -третья степень. Некоторые растущие в дождевом лесу деревья вообще лишены ветвей (пальмы, древовидные папоротники, некоторые двудольные), но зато у них развиты особо крупные листья, или вайи.
Одна из самых характерных черт влажнотропических деревьев - досковидные и ходульные корни. Досковидные выросты у основания ствола чаще всего встречаются у деревьев яруса А. Несомненно, что это приспособление для дополнительной опоры, ибо общее проникновение корней в почву у деревьев дождевого леса крайне мало и часто не превышает 0,5 м. Ходульные корни встречаются значительно реже, они более характерны для болотистых местообитаний (а также в манграх) и свойственны преимущественно низкорослым видам из яруса С. Как досковидные, так и ходульные корни встречаются у видов из разных семейств. Следовательно, это признак, отражающий в первую очередь сходные экологические условия местообитания. У некоторых видов наличие или отсутствие ходульных корней зависит от места произрастания. Например, в Уганде у музанги (Musanga cecropioides) таких корней нет, а в Западной Африке всегда есть. Деревья с досковидными корнями чаще всего встречаются в самых теплых и постоянно влажных районах, хотя отдельные представители проникают и в дождезеленые тропические леса. В постоянно влажных, но прохладных горных лесах тропиков (нефелогилеях) деревья с досковидными корнями исчезают. На почвах с хорошим дренажем таких деревьев также намного меньше, поскольку в таких местах корневая система проникает значительно глубже.
Мы уже отмечали, что однообразие внешнего вида деревьев влажной гилей во многом обусловлено сходством внешнего вида листьев у самых разнообразных видов. Более того, листва одного и того же яруса особенно однообразна, поэтому у одного и того же дерева листья с верхних и с нижних ветвей могут различаться, а у разных деревьев листья из одного яруса обычно почти неразличимы. По форме и жесткости большинство листьев похожи на листья всем известного фикуса, растущего у нас в комнатах (Ficus elastica); характерна капельница - вытянутый носик на конце листа. В нижних ярусах размер листьев примерно такой же, как и у фикуса; в верхних ярусах листья мельчают и более напоминают лимон или лавровишню, размер капельницы уменьшается. Перистые листья некоторых бобовых-; а также характерные листья пальм, драцен или панданусов сравнительно редко разнообразят общий фон. Они значительно чаще определяют облик растительности во вторичных или в прибрежных и болотных участках.
Одна из самых примечательных картин во влажных тропиках - распускающиеся молодые листья. По окраске они необычайно разнообразны и резко отличаются от общего темно-зеленого фона - листья бывают ярко-малиновые, бордовые, почти белые, желтые, бледно-зеленые, синевато-стальные. Как писал П. Ричарде, «яркость окраски листвы часто может обмануть даже опытного наблюдателя, заставив его думать, что деревья находятся в цвету». Молодые листья и целые побеги вначале висят, как увядшие, а через одну-две недели начинают зеленеть, становятся жестче и постепенно принимают обычное положение. Цветные и поникшие молодые листья и побеги называют листьями-платками. Такие листья распускаются очень быстро, иногда за одну ночь, они как бы «высыпают» на дереве.
Причина вялости молодых побегов - разрыв во времени между началом роста и началом дифференцировки тканей. Распускание почек идет за счет клеток меристемы и формирование механической ткани отстает. В иных климатических условиях более жесткий контроль среды не дает существовать такому «излишеству» - молодые побеги в случае недоразвитости механической ткани просто засыхали бы. Кстати, во время относительно сухих периодов усыхание листьев-платков отмечено и в пределах влажных тропиков. Иногда вялость и окраску листьев-платков рассматривают в качестве приспособительных признаков. Однако П. Ричарде считает, что говорить в данном случае о приспособительных реакциях вряд ли стоит: «Задержка в приобретении тканями жесткости, которая могла бы стать вредоносной для растения в более суровой среде, здесь остается «безнаказанной»... С этой точки зрения можно расценивать и многие другие признаки растений тропического дождевого леса, считающиеся приспособительными»1.
Читатель легко может получить представление о листьях-платках, об их быстром росте, цвете и поникшем виде, не выезжая в тропики. Для этого достаточно у зрелого плода манго, которые нередко продают у нас во многих городах, вынуть семечко, а затем без долгого хранения посадить его в легкую землю и поддерживать постоянную влажность и высокую температуру (днем при +30...+35°С). Такие условия можно создать летом в оконной теплице, которую легко соорудить из аквариума или из натянутой на каркас полиэтиленовой пленки. Росток манго быстро вытягивается и образует ярко-бордовые блестящие листья, которые свисают вниз, но очень быстро увеличиваются в размерах. После того как листья вырастают, они начинают зеленеть, в основании черешка формируются чрезвычайно характерные для листьев влажных тропических деревьев утолщения, листья принимают обычное положение.
Интенсивность транспирации и фотосинтеза древесных растений вечнозеленых тропических лесов колеблется весьма значительно и на единицу массы в общем укладывается в те же пределы, что известны и для умеренных широт. Характерны относительно низкие величины транспирации в полуденные солнечные часы. Это объясняется перегревом освещенных листьев, температура которых может превышать температуру воздуха на 10-15°С. Избыточные температуры становятся все более опасными при повышении температуры воздуха, ибо у поверхности листа градиент давления водяного пара повышается быстрее, чем градиент температуры. Для устранения чрезмерной потери воды необходима толстая кутикула, препятствующая высыханию и гибели листьев при закрытых устьицах (при этом поверхность листа особенно нагревается). «Следовательно, деревьям тропического дождевого леса в течение нескольких часов приходится испытывать недостаток влаги, по-видимому не менее жесткий, чем в аридных областях. Именно этими условиями следует объяснить часто встречающуюся ксероморфную структуру листьев, благодаря которой растения получают возможность постоянно поддерживать стабильный водный баланс с высокой гидратурой (низкое осмотическое давление), несмотря на временами большой дефицит влажности листьев»2. Приведенная цитата показывает очень характерную и не всегда учитываемую черту влажной гилей. Кратковременные дефициты влажности обычны у деревьев яруса А. Именно в этом ярусе листья чаще всего имеют кожистую, явно ксероморфную структуру. Такую же структуру имеют и эпифиты верхних ярусов леса.
Образование цветков и плодов на стволах или на толстых деревянистых ветвях - явление каулифлории - хорошо известная особенность древесных растений (деревьев, древесных лиан) влажных тропиков. Считают, что во всем мире более 1000 видов способны к каулифлории. За пределами влажных тропических лесов лишь единицы деревьев и кустарников обладают такой особенностью. Существует множество попыток объяснить каулифлорию с позиций приспособляемости к условиям среды (климатические особенности, приспособления к опылению или к распространению семян, защита плодов и цветков от поедания и т. п.), однако убедительной и стройной картины пока нет. Необходимы дополнительные экспериментальные материалы и их разносторонняя биоценологическая интерпретация. Громадное большинство видов, обладающих каулифлорией, приурочено к ярусу С или даже D. Интересно, что именно ярус С чаще всего оказывается наиболее сомкнутым, именно по кронам этого яруса пролегают пути многочисленных древесных позвоночных - зверей и птиц, потенциальных потребителей семян и плодов. В то же время цветки, расположенные на стволах под сомкнутым пологом яруса С, оказываются более доступными для опылителей.
В самых нижних ярусах влажных тропических лесов -в ярусах D и Е преобладают молодые деревья, с возрастом переходящие в вышерасположенные ярусы. В отличие от наших лесов, разнообразие и обилие трав под пологом леса невелико. Вообще ярусы Е и D очень часто оказываются фрагментарными, разорванными на отдельные сравнительно небольшие куртины. В первую очередь это связано с недостатком света и с корневой конкуренцией. В тропиках у трав надземные органы живут длительное время, характерны также отдельные высокорослые растения - бананы (Musa) и другие представители порядка имбирных (Zingiberales - Maranta, Ravenala и т. п.). Особенно высокие (до 6 м) представители этих трав уже входят в ярус С.
В составе ярусов Е и D преобладают немногие виды однодольных и двудольных, а также папоротники и селагинеллы. Несмотря на значительное разнообразие внешнего вида лесных трав нижнего яруса, проявляется конвергентное сходство некоторых представителей разных систематических групп. Особенно удивительны широкие листья злаков и осоковых. Большинство трав является фанерофитами, а не гемикриптофитами или геофитами, как в более изменчивых условиях тепла и влаги.
Некоторые травы обладают пестро окрашенными листьями, более привлекательными, нежели обычно невзрачные цветки. Последние нередко развиваются на коротких цветоносах, расположенных у самой поверхности земли. В условиях очень высокой влажности, царящей под пологом гилей, травы транспирируют весьма слабо. Зато многие из них способны избавляться от влаги путем гуттации. Чрезвычайно низкая интенсивность фотосинтеза в условиях затенения компенсируется столь же низкой интенсивностью дыхания. В результате баланс оказывается положительным и растение накапливает массу.
Обычная для умеренных широт жизненная форма кустарников во влажной гилее практически отсутствует и заменяется карликовыми деревьями. В небольшом числе встречаются частично одревесневающие полукустарники.
Обращает на себя внимание обязательное присутствие лиан -вьющихся и лазающих растений, опорой для которых служат деревья. Большинство лиан имеет одревесневающий ствол, длина которого может превышать 100 м, хотя чаще всего остается в пределах нескольких десятков метров. Большинство лиан достигает яруса В. Особенно много их в ярусе С. Именно благодаря лианам в лесу заполняются все сколько-нибудь заметные «пробелы» яруса С. Листья лиан сходны с листьями соответствующего яруса древостоя, но по форме более компактны и округлы. Хорошим примером такой лианы может служить сциндапсус, который часто разводят в комнатах.
Между лианами и другими экологическими группами зеленых растений тропических лесов существуют постепенные переходы. В частности, выделяют такую промежуточную между лианами и эпифитами группу, как полуэпифиты. Последние развиваются вначале как лианы, но после закрепления на стволе или ветвях дерева-хозяина основной стебель засыхает и растение переходит к эпифитной жизни. Нередко наблюдается обратное. Эпифитное вначале растение пускает вниз воздушные корни, которые укореняются, одновременно опутывая хозяина. Дальнейшее развитие последней тенденции демонстрируют растения-удушители, которые после прорастания и недолгой эпифитной стадии опускают к почве многочисленные воздушные корни, которые срастаются между собой, опутывают растение-хозяина, утолщаются и постепенно душат дерево-хозяина. Это дерево гибнет, а на его месте остается уже выросшее другое дерево. Классическими примерами растений-удушителей являются различные фикусы, но эта жизненная форма встречается нередко и среди арендных, а также и других семейств. Однако далеко не всегда удушители полностью губят хозяина. Промежуточной группой между паразитами и эпифитами можно считать полупаразитов, которые самостоятельно фотосинтезируют, но минеральные питательные вещества и воду получают из побегов хозяина. В тропических лесах, как и на юге Европы, в эту группу входят виды омелы - представители семейства Loranthaceae. В каждом районе встречается немало родов и множество видов этого семейства.
Наконец, крайне разнообразны представители группы эпифитов, без которых невозможно себе представить тропическую гилею. Эпифиты растут на стволах и ветвях других растений (чаще - деревьев), используя хозяина только как субстрат для опоры. Выделяют также группу эпифиллов, которые поселяются на поверхности листьев. Эпифиллы представлены главным образом водорослями, лишайниками и мхами. Эпифиты также насчитывают много мхов и лишайников, однако наиболее заметны и разнообразны эпифиты сосудистые, т. е. папоротникообразные и цветковые, они доминируют по массе. Поселяясь на стволах и ветвях деревьев, эпифиты выигрывают в освещенности и избегают корневой конкуренции в почве, однако вынуждены приспосабливаться к более жесткому режиму увлажнения, к недостатку субстрата для извлечения минеральных солей. Все это приводит к поразительному разнообразию адаптации, подробно описать большинство из которых, к сожалению, здесь невозможно. Можно, однако, указать на такие интересные черты, как мирмекофильность многих эпифитов (симбиоз с муравьями), формирование «висячих аквариумов» из плотных розеток листьев бромелий, специальные образования для удержания на коре хозяина (листья-скобки), клетки и чешуйки для поглощения воды.
Разделение ризосферы и филлосферы условно, так как из-за высокой влажности воздуха множество корней деревьев и, разумеется, эпифитов развивается над поверхностью почвы. Впрочем, эпифиты формируют своеобразный висячий ярус почв, чему способствуют термиты, муравьи и другие животные, а также опад с деревьев.
Весьма примечательно, что даже в самых влажных вариантах вечнозеленого дождевого леса равнин могут присутствовать суккуленты - растения с мясистой водозапасающей тканью. И суккуленты появляются именно в форме эпифитов. Таковы, к примеру, рипсалисы из кактусовых (Rhipsalis), некоторые бромелиевые, ластовневые. В гилеях с более или менее выраженным относительно сухим периодом число суккулентов среди эпифитов повышается. Особенно это заметно в Новом Свете с его огромным разнообразием кактусовых и бромелиевых. В пределах вечнозеленых лесов весьма обычны такие эпифитные кактусовые, как виды родов Epiphyllum, Epiphyllopsis, Epiphyllanthus, Zygocactus, Schlumbergera, Lepismium, Aporocactus, Selenicereus, Disocactus, Hatiora, Hylocereus, Marniera, Mediocactus, Pseudorhipsalis, Pseudozygocactus, Rhipsalidopsis, Strophocactus, Weberocereus. Указанные 19 родов эпифитных кактусовых - заметная доля от всего списка из 230 родов этих растений. Виды этих 19 родов встречаются частично и в пределах дождезеленых лесов, иногда они могут расти также на поверхности скал и просто на поверхности земли. Последнее, однако, встречается уже за пределами вечнозеленых лесов, в их же пределах виды отмеченных родов всегда ведут эпифитный образ жизни.
У эпифитных кактусов (это свойственно почти всем представителям семейства), а также у многих эпифитных орхидей и бромелий отмечен так называемый «эффект Соссюра», типичный и для большинства суккулентов. Сущность его состоит в том, что днем на свету растение замыкает устьица и прекращает транспирацию (и потерю воды). Однако закрытые устьица препятствуют также газообмену, необходимому для фотосинтеза. Растения разрешают это противоречие. Ночью устьица открыты, происходит поглощение углекислоты и связывание ее в клеточном соке в виде органических кислот. К утру кислотность сока максимальна.На свету же углекислота вовлекается в фотосинтез, который происходит, таким образом, «за закрытыми дверями». Среди эпифитов мы находим множество красиво цветущих растений, и непревзойденными в этом отношении являются орхидные. Красивые цветки свойственны также видам геснериевых (например, из рода Columnea).
Одна из самых замечательных особенностей дождевого леса -абсолютное флористическое господство древесных растений. Большая часть видового списка как большого лесного района, так и пробной площадки в 1-2 га приходится на деревья. В составе древостоя чаще всего не бывает резко доминирующих на большой площади одного-двух видов, преобладают полидоминантные группировки. Даже если выявляется один или несколько наиболее многочисленных видов деревьев, то они сопровождаются десятками других видов. Так, например, в южной части провинции Юннань (КНР) на площади в 0,25 га (50 на 50 м) А.Г. Вороновым было отмечено 62 вида деревьев. В травяном покрове присутствовало только 20 видов. Более 60 экземпляров на этой площади имели только два самых массовых вида - Knema wangii, Gironniera subaequalis, более 10 экземпляров имели 7 видов, а до 10 экземпляров - 54 вида.
Исходя из количества фоновых видов и характера доминирования, влажный экваториальный лес представляет собой как бы красочный луг умеренных широт, где травы заменены деревьями. И наоборот, богатый видами луг умеренной полосы (или некоторые варианты красочных степей) представляет собой как бы экваториальную гилею, в которой древесные растения заменены травами. Указанная аналогия не только удобна для уяснения особенностей растительного покрова влажных экваториальных лесов, но имеет и более глубокий смысл. Дело в том, что исторически (филогенетически) травы — более молодая жизненная форма. В то же время влажная гилея - наиболее древний из сохранившихся типов формаций. Она в той или иной степени была исходной для образования растительности более высоких широт. Многие семейства и даже роды, представленные в умеренных широтах лишь в виде трав, в дождевом лесу на экваторе или у тропиков - настоящие деревья, порой очень высокие, или же толстые деревянистые лианы.
Древесные растения преобладают также в горных лесах влажных тропических регионов. Древесные виды из групп, представленных в умеренных широтах травами, находят и на некоторых островах южного полушария (например, на острове Св. Елены). Наш замечательный ботанико-географ А.Н. Краснов так описывал свои впечатления от восхождения на вершину горы Геде на Яве в 1892 г.: «На смену тропической флоре приходит альпийская, и всего любопытнее то, что, как мне всюду приходилось видеть, эти немногие, дошедшие до вершины формы слагаются в ассоциации, большей частью представленные теми же родами, которые характеризуют нашу тундру далекого севера. Разница, <
Дата добавления: 2019-12-09; просмотров: 1511;
Поиск по сайту
Узнать еще
- Автомобили-тягачи для вывозки леса
- Анализ процесса качения эластичного колеса по деформируемому грунту
- Антропогенные воздействия на леса и другие растительные сообщества
- Б. Дипольные электрические зондирования: экваториальные, азимутальные и осевые
- Бесстензельная перевозка леса.
- Биологический круговорот масс химических элементов в фитоценозе дубового леса Северной Венгрии
- В продуктах опада деревьев-эдификаторов леса Уокер Бранч,
- Введение в теорию качения колеса

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
