ПО АНАЛОГИЧНЫМ ПРИЗНАКАМ
Для классификации биоценозов используют уже разработанные в фитоценологии схемы, поскольку растительность выступает наиболее удобным индикатором всего биотического сообщества. Таких схем существует довольно много, но обычно самой мелкой единицей признают ассоциацию (тип фитоценоза). Ассоциация вместе с соответствующими ей абиотическими компонентами соответствует самой мелкой единице классификации биогеоценозов - типу биогеоценоза. Ассоциация характеризуется одними видами - доминантами - каждого яруса, сходной пространственной структурой и условиями существования. Следующие за ассоциацией таксономические категории включают группы ассоциаций, формации, группы и классы, формаций, тип формации (тип растительности).
Вплоть до уровня формаций главный критерий - систематическая общность каких-либо элементов сообществ (для формаций - общность видов-эдификаторов, т. е. доминирующих видов с выраженной средообразующей способностью). Следовательно, здесь существенны гомологичные признаки сообществ. В некоторых схемах и более обобщенные таксоны, объединяющие разные формации, также группируются в значительной степени по гомологичным признакам (например, флороценотип). В то же время можно считать, что на низших ступенях классификационной системы подходы аналогичный и гомологичный практически совпадают (близкие или тождественные виды, соответственно, чаще всего представляют близкие или тождественные жизненные формы).
Обычно в одну группу формаций объединяют те из них, в которых эдификаторы относятся к сходным жизненным формам. Например, выделяют такие группы формаций, как темнохвойные леса, светлохвойные листопадные леса, широколиственные леса, крупнозлаковые луга, мелкозлаковые луга и т. п. Среди классов формаций приводят в качестве примеров хвойные леса, листопадные летнезеленые леса (сбрасывающие листву в холодный сезон), листопадные дождезеленые леса (сбрасывающие листву в сухой сезон) и т. п. Понятие о классе и типе формации близко к понятию «биом». Под биомом при этом чаще всего понимают зональный класс или тип формации, свойственный региону с данными климатическими особенностями. Территориально, картографически классы и типы формаций и биомы совпадают с системой физико-географических, или природных, зон.
Легко видеть, что в классификационных категориях от группы формаций до типа формаций включительно господствует принцип сходства по аналогии - обычно по эколого-морфологическим особенностям эдификаторных видов растений. На самом же деле под прямым и косвенным влиянием сходных условий среды глубокие аналогии наблюдаются и в других характеристиках сообществ - в уровне первичной и вторичной продукции, в соотношениях биомасс и разнообразии разных структурных и функциональных элементов, по геохимическому режиму и т. п. В то же время некоторые адаптации к сходным условиям могут быть реализованы разными способами, не говоря уже о том, что в разных регионах - средствами различных флор и фаун. Подробнее особенности гомологически различных биоценозов-аналогов освещены в специальной части при разборе зональных типов сообществ (в ранге класса и типа формации).
Картографическое отображение классов формации вполне допустимо даже в мелком масштабе (т. е. мельче миллионного). В качестве примера можно привести карту распределения первичной продукции и биомассы (с приростом ее за год) в масштабе 1:110 000 000 (рис. 10, а и 10, б). Именно с позиции такого масштаба (близкого к стомиллионному) в специальной части будут рассмотрены описания основных биомов суши. Но и при столь высокой степени генерализации мы вынуждены уделить внимание лишь ключевым типам сообществ, упомянув остальные весьма бегло. Распределение именно таких основных биотических сообществ, выделенных по аналогичным признакам, представлено на приводимой карте. Соответствующие тем или иным выделам разделы специальной части можно также рассматривать в какой-то степени в качестве развернутого пояснения (легенды) к этой карте.
Каждый выдел очерчивает регион (индивидуально-хорологическое подразделение) на основе аналогичных характеристик, поэтому упомянутая карта одновременно может служить основой биогеографического районирования по таким характеристикам.
Если бы не существовало преград к расселению, то филогенетическая структура внутри каждого из них оказалась бы выровненной или даже одинаковой. В этом случае результаты аналогичного и гомологичного подходов полностью совпали бы. Иными словами, представленные выделы показывают в принципе возможные (но только по гигротермическим требованиям) потенциальные ареалы для видов, населяющих хотя бы малую часть одного выдела. К сожалению, процесс бессистемной и даже неосознанной интродукции уже привел в ряде случаев к расширению ареалов некоторых видов до потенциально возможных по условиям среды. К сожалению потому, что такого рода «обогащение» почти неминуемо ведет к возникновению сильной конкуренции и к вытеснению более «слабых» видов. В результате в масштабе биосферы генетическое разнообразие падает.
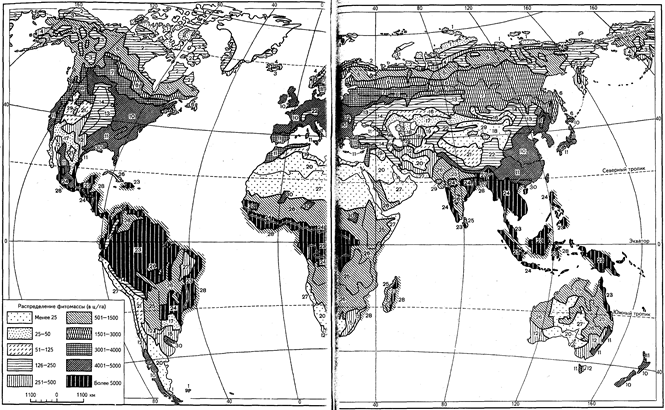
Рис.10, а. Распределение надземной и подземной фитомассы суши
(в ц/га сухого органического вещества):

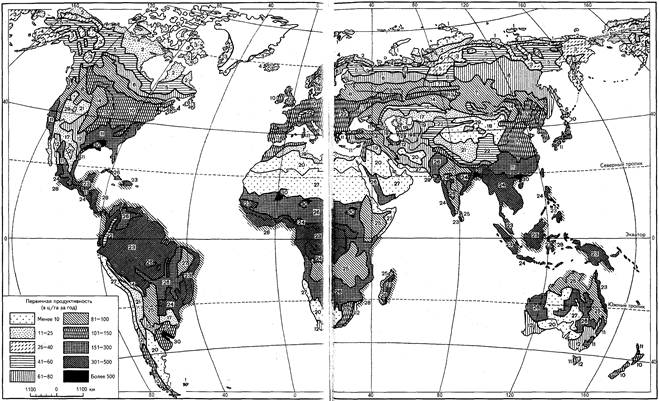
Рис.10, б. Распределение первичной продуктивности, или общей продуктивности, биоценозов суши (в ц/га за год сухой фитомассы):

Дата добавления: 2019-12-09; просмотров: 933;
Поиск по сайту
Узнать еще
- Виды цен и их классификация по различным признакам.
- Волны н a водных объектах классифицируются по различным признакам.
- Другие показатели наличия связи между признаками.
- Классификация грузов по различным признакам
- Классификация затрат, формирующих себестоимость продукции, по различным признакам.
- Классификация зданий и помещений по признакам пожарной опасности
- КЛАССИФИКАЦИЯ ИНФОРМАЦИИ ПО РАЗНЫМ ПРИЗНАКАМ
- Классификация оборудования по монтажным признакам

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
