И САВАННОВЫЕ РЕДКОЛЕСЬЯ
Географическое распространение, гигротермический
и геохимический режимы
К северу и югу от зоны вечнозеленых экваториальных лесов в пределах тропических широт распространены крайне разнообразные сообщества, приспособленные к перенесению сухого периода года. В этот период температуры остаются высокими, однако увлажнение оказывается недостаточным для бесперебойной вегетации растительности. Остальное время идут дожди, и влажность нередко оказывается даже избыточной. Следовательно, влажный период здесь сходен с условиями, характерными круглый год для настоящих тропических и экваториальных дождевых лесов.
Сезонная засуха может быть выражена в разной степени в зависимости от продолжительности периода дождей и от общей суммы выпавших за год осадков. Очень большое влияние на облик сообществ оказывают также местные эдафические условия и деятельность человека. Приведенный на рисунке 18 ряд климадиаграмм иллюстрирует главную особенность гигротермических условий таких регионов - высокие температуры в течение всего года и резкую сезонность увлажнения.
На приведенных профильных диаграммах (рис. 19) можно проследить, как по мере усиления сезонной засухи вечнозеленый дождевой лес сменяется вечнозеленым сезонным лесом, полувечнозеленым лесом, сухим листопадным (дождезеленым лесом) и далее еще более ксерофильными сообществами. Приведенные схемы показывают, что увеличение засушливости в тропиках отнюдь не обязательно приводит к появлению открытых участков, поросших злаками, среди которых растут отдельные куртины деревьев. Обширные злаковники с редкими вкраплениями древесных растений (высокотравные и типичные саванны) чаще всего возникают как результат регулярного выжигания сухой травы или приурочены к особым почвенно-гидрологическим условиям (чередование иссушения и избыточного увлажнения с застаиванием воды в сезон дождей). Как уже отмечалось в предыдущем разделе, вторичные саванны могут возникать даже в пределах зоны влажных гилей после сведения леса и последующего регулярного выжигания травы.
В то же время разреженные древостой из листопадных пород, в приземном ярусе которых господствуют злаки, можно рассматривать в качестве коренных сообществ. Такие сообщества, как правило, характерны для районов с годовой нормой осадков менее 800 мм. В Австралии более распространены редколесья из вечнозеленых видов эвкалиптов, а также филлодийных акаций. Бразильские редколесные группировки, так называемые «кампос серрадос» (campos cerrados - закрытые поля, заросшие открытые пространства), также в значительной степени содержат вечнозеленые формы деревьев и кустарников. Однако эти сообщества часто произрастают в более увлажненных районах, с осадками свыше 1000 мм в год. Часть их, безусловно, представляет собой стадии регрессивной сукцессии на месте сомкнутых лесов.
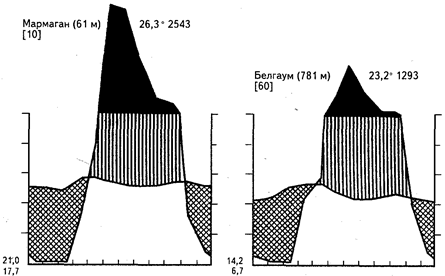
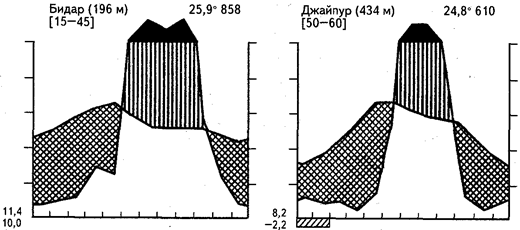
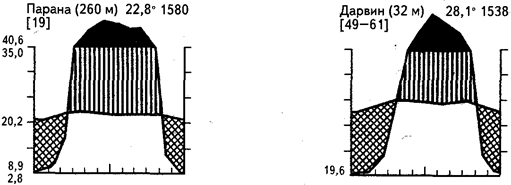
Рис. 18. Климадиаграммы тропических переменно-влажных районов
с летними дождями и зимней засухой
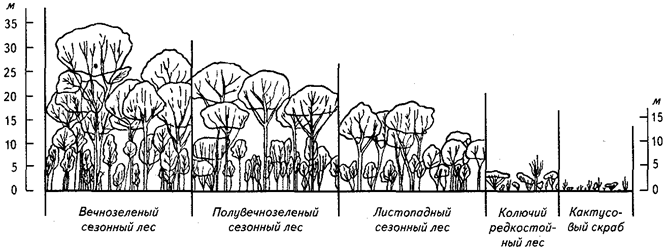
Рис. 19. Профильные диаграммы различных классов лесных формаций
на низменностях острова Тринидад и прилегающих частей Венесуэлы
(по П. Ричардсу, 1961)
Резко выраженная сезонность физических факторов среды приводит к соответствующей сезонности геохимических процессов. В период дождей господствует промывной режим почв, во время засухи почвы иссушаются и происходит подтягивание солей к поверхности. В то же время, в отличие от аридных и семиаридных группировок, засоление поверхностных горизонтов нехарактерно. Оно может проявляться только в наиболее засушливых районах, периодически заливаемых дождями и имеющих плохой дренаж из-за материнских пород тяжелого механического состава. В открытых саваннах почвы обеднены гумусом по сравнению с лесами, развитыми в тех же климатических условиях. Периодическое переувлажнение и иссушение приводит также к образованию на небольшой глубине уплотненного железистого горизонта красного цвета, который при эрозии часто выходит на поверхность. Такие почвы бедны азотом и фосфором. В районе развития кампос серрадос Бразилии отмечалось также обеднение почвы некоторыми микроэлементами (бор и цинк), калием и фосфором. Одновременно с этим почва может содержать чрезмерное количество алюминия, что угнетающе действует на многие древесные растения.
Сообщества фотоавтотрофов
Группировки фотоавтотрофов в тропических сезонно-засушливых районах чаще всего формируют редкостойные древостой, между которыми развит злаковый покров или иногда колючие и суккулентные кустарники. Очень часто формируются также большие пространства злаковников с отдельными группами деревьев, устойчивых к огню. На большей части территории периодические палы сухого травостоя оказывают мощное воздействие на облик растительности. Коренные сообщества в виде светлых и сомкнутых листопадных (дождезеленых) лесов сохранились в меньшей степени. Леса чаще всего заменены производными саваннами.
По степени увлажнения выделяют три группы растительных сообществ: 1) самый влажный пояс с тропическими влажными дождезелеными (муссонными - в Азии) лесами, редколесьями типа кампос серрадос Бразилии, высокозлаковыми (влажными) саваннами; 2) пояс средней влажности с листопадными (дождезелеными) сухими лесами, колючими лесами и саваннами с злаковым покровом средней или небольшой высоты, без вечнозеленых галерейных лесов; 3) колючие и суккулентные кустарники и леса типа «каатинга» (что в переводе означает «белый лес») на северо-западе Бразилии, сухие злаковники с колючими кустарниками. Два первых пояса примерно соответствуют гвинейской и суданской саванне в Африке, которые образуют хорошо выраженные зональные массивы к северу и востоку от вечнозеленых лесов. К югу от этих лесов расположены светлые редкостойные леса миомбо, которые при нарастании сухости сменяются саванновыми редколесьями.
В рамках этой книги невозможно описать все разнообразие тропических сезонно-сухих биоценозов. Поэтому главное внимание сосредоточено лишь на некоторых коренных типах леса, а затем на саваннах и редколесьях, занимающих большие площади, но часто возникших лишь под влиянием деятельности человека.
Наиболее сухие типы тропического дождевого леса - сезонные вечнозеленые леса. Эти леса испытывают относительно короткий период засухи (до 2-3 месяцев), однако из-за большой суммы годовых осадков (свыше 2000 мм) влаги в почве оказывается достаточно для деревьев, которые остаются зелеными. Прерывают вегетацию некоторые эпифиты и травянистые растения. При увеличении сухости самые крупные деревья сбрасывают на короткое время листву, нижние же ярусы (В и С) никогда не оголяются одновременно (см. рис. 19). Между такими типами леса и вторичными саваннами нет резкой разницы, и они могут встречаться даже в одном районе, занимая несколько различные местообитания. Так, в Заире, к юго-западу от Киншасы, на фоне преобладания вторичных саванн попадаются массивы старых вторичных полувечнозеленых и даже сезонных вечнозеленых лесов (последние приурочены к понижениям). В последних числах сентября, в самом конце сухого сезона, который длится четыре месяца, подавляющее большинство деревьев остаются зелеными, многие деревья и лианы цветут, однако апифитные орхидеи в нижнем древесном ярусе пребывают в покое, а под ногами шуршит и местами даже пылит сухая листва. На прогалинах и опушках злаковый покров совершенно сухой, но некоторые геофиты (кринум из амариллисовых) уже трогаются в рост и выбрасывают цветочную стрелку и свежие зеленые листья.
При ухудшении увлажненности начинают преобладать листопадные древесные виды. Листопадные дождезеленые леса в Африке шире всего представлены на юге материка - в Замбии, Анголе, Мозамбике, Заире. Эти леса образованы главным образом невысокими (около 10-12 м) ветвистыми деревьями с перистыми листьями из семейства бобовых. Часто отчетливо доминируют немногие виды из родов брахистегия (Brachystegia), изоберлиния (Isoberlinia-Julbernardia), весьма обычны также некоторые виды акаций и альбицций. Сомкнутость крон обычно составляет 0,6-0,8. Под пологом таких светлых лесов хорошо развит кустарниковый ярус, злаковый покров занимает прогалины и опушки. Часто прогалины образуются в результате деятельности слонов. Эти леса (миомбо) легко повреждаются огнем и переходят в производные саванновые сообщества.
К северу от бассейна Конго (Заира) дождезеленые леса сохранились лишь отдельными массивами среди обширных саванн. Непосредственно к дождевому лесу примыкают вторичные высокотравные саванны и редколесья. Здесь встречаются многочисленные представители настоящего дождевого леса, способные переносить выжигание трав (например, масличная пальма Elaeis guinensis). Еще севернее расположена полоса так называемых гвинейских саванн, которые также характеризуются мощным высокотравьем и редкими древостоями из выносливых к огню Daniella oliveri, Lophira alata, Terminalia glaucescens. В немногих местах редколесья становятся все более и более густыми, включающими уже настоящие лесные дождезеленые виды, плохо переносящие выжигание травы. Лишь малыми клочками еще встречаются настоящие леса, в которых представлены древостой из Khaya grandifolia, Triplochiton scleroxylon, Piptadenia. В несколько более засушливых районах (но имеющих годовую сумму осадков более 800 мм) редколесные и лесные группировки в первую очередь состоят из Isoberlinia doka, I. tomentosa, Monotes kerstingii, Uapaca somon, T. glaucescens, Anogeissus schimperi, Malacantha ainifolia, Khaya senegalensis, Afzelia africana, Diospyros mespiliformis, видов Pterocarpus, Strychrios, Sclerocarya, Burkea, Butyrospermum parkii.
Виды Isoberlinia и Anogeissus schiperi местами образуют монодоминантные древостой.
На восток от бассейна Конго (Заира) в приэкваториальных регионах с относительно сухим климатом бывают два периода дождей. Это способствует формированию колючих зарослей с обилием суккулентов, в первую очередь древовидных молочаев, характерных для рифтовой долины севернее озера Киву и более восточных территорий.
Более засушливая полоса суданских саванн представлена главным образом регулярно выжигаемыми злаковниками, на фоне которых растут отдельные баобабы (Adansonia digitata) и группы небольших зонтиковидных деревьев и кустарников (Acacia ssp., Combretum glutinosum, Balanites aegyptiaca, Lannea microcarpa, Sclerocarya birrea).
Только на каменистых возвышенностях травяной покров слабо развит, ежегодные палы не повреждают деревьев и образуются группировки дождезеленых листопадных лесов с господством Lonchocarpus, Stereospermum, Sterculia, Anogeissus, Boswellia, Ficus. В то же время в южной половине Африки леса миомбо переходят во все более ксерофитное редколесье мопане из акаций, альбицций и дерева мопане (Colophospermum-Copaifera mopane). Здесь также встречаются баобаб, коммифора и заросли сухих кустарников. Леса мопане имеют более развитый злаковый покров,
Между суданской саванной и Сахарой протянулась переходная зона под названием Сахель, или сахельская саванна («сахель» по-арабски означает «берег», к которому стремились, пересекая караванами пустыню). Сахельская саванна представляет собой разреженное акациевое редколесье с очень скудным злаковым покровом из однолетних видов. Этот район в течение столетий интенсивно используют под пастбища, а древесные растения вырубают для топлива. Флористически Сахель резко отличен от лежащих южнее территорий, и, видимо, его лучше рассматривать вместе с пустынными группировками Сахары. На юге Африки также распространены разреженные акациевые саванны, переходящие в пустыню.
Злаковый покров в наиболее увлажненных районах Африки формируется императой (Imperata cylindrica) или слоновой травой (Pennisetum purpureum), которая может достигать нескольких метров в высоту, напоминая плантацию сахарного тростника. В типичных саваннах разных материков чаще всего доминируют виды бородача (Andropogon), аристиды (Aristida), гипаррении (Hyparrhenia), темеды (Themeda). Для Австралии очень характерны триодии (Triodia). Нередко тропические злаковники включают также виды ковылей, овсяниц, вейников, хотя чаще эти злаки приурочены к поднятиям. Во вторичных злаковниках Азии также обычна императа, очень часто встречается дикий сахарный тростник (Saccharum spontaneum). В общем можно заключить, что на уровне родов, а иногда даже и видов злаки распространены значительно шире, нежели древесно-кустарниковые компоненты саванновых или редколесных сообществ. Это широкое распространение выражается как в общности родов злаков в тропических широтах разных материков, так и в проникновении многих этих родов в умеренные широты. Для тропических саванн типично преобладание многолетних злаков. Частое выжигание и пастбищная перегрузка ведут ко все более заметному участию однолетних злаков.
Дождезеленые тропические леса хорошо представлены в Евразии. Обычно их называют муссонными лесами, выделяя влажные и сухие варианты. В Индии и прилегающих странах влажные муссонные леса образованы преимущественно тиком, или тэком (Tectona grandis), и салом (Shorea robusta), а также ксилией (Xylia xylocarpa). Более сухой вариант включает, кроме тика, Terminalia tomentosa и некоторые виды Pentacme, Diospyros и Dypterocarpus. В районах с годовой нормой осадков менее 1000 мм отсутствуют высокоствольные леса и коренные сообщества представлены листопадными и колючими редколесьями из различных акаций.
Дождезеленые тропические леса Евразии, как и вечнозеленые, очень часто образованы видами семейства диптерокарповых (Dipterocarpaceae), большая часть которых сосредоточена в Азии и немногие встречаются также в Африке. Так, в Азии и на Новой Гвинее произрастает около 100 видов из рода Shorea, около 70 видов из родов Hopea, Dipterocarpus, Pentacme, Vateria. Тик относится к вербеновым, большую роль играют также представители бобовых.
В Австралии листопадные леса почти не представлены. Тропические сезонно-засушливые области заняты эвкалиптовыми и акациевыми редколесьями. В сухой сезон хорошо развитый злаковый покров высыхает, но эвкалипты сохраняют зеленую листву. По мере дальнейшей аридизации все более заметны акации с филлодиями, т. е. с расширенными черешками листьев, выполняющими функцию фотосинтеза. Ландшафтными становятся также казуарины (Casuarma) с зелеными молодыми побегами и редуцированными крохотными листочками. Побеги эти фотосинтезируют. Внешне они похожи на хвою сосны.
Сходство с хвойными дополняют своеобразные «шишки» казуарин. Однако эти растения принадлежат к особому порядку и семейству двудольных. На северо-востоке материка сухие акациевые редколесья переходят в группировки, которые характеризуются присутствием различных низкорослых деревьев с вздутыми толстыми стволами (брахихитон - Brachychiton australe из семейства стеркулиевых). Под пологом деревьев густо разрастается кустарниковый ярус, злакового покрова нет. Большинство деревьев листопад но, среди кустарников около половины экземпляров принадлежит вечнозеленым видам.
В разных районах Центральной и Южной Америки очень богатый набор сезонно-сухих сообществ. Чаще всего в средних эдафических условиях можно наблюдать постепенный переход от вечнозеленых лесов к пустыням. При этом комплексы древесных растений со злаковниками и другими травами (осоковые) могут выпадать и заменяться комплексами древесных растений с колючими кустарниками и суккулентами. Занимающие обширную площадь в долине Ориноко злаковники (низкие льянос) с отдельными куртинами маврикиевой пальмы (Mauritia flexuosa) образовались под влиянием периодического затопления и иссушения почвы. Впоследствии эти злаковники расширили свой ареал и за счет выжигания травы во время засухи. Влияние частых палов отчетливо видно и в области кампос серрадос в Бразилии. Нередко встречаются и своеобразные эдафические условия из-за каменистости грунтов. Здесь обильны суккуленты и некоторые древесные растения. Остановимся подробнее на описаниях нескольких растительных сообществ Южной Америки.
Большую часть Бразильского нагорья занимают группировки кампос серрадос. Они довольно разнообразны даже внешне, хотя чаще всего это низкорослое редколесье или саванно-видная растительность с хорошо развитым злаковым ярусом. Осадки выпадают летом, общая годовая норма весьма значительна (от 1200 мм и более). Характерна мощная кора выветривания (до 20 м), а также постоянно влажный горизонт глубже 2 м, доступный корням многих деревьев и кустарников, но недоступный злакам. Последние в сухой период года высыхают (преобладают виды из родов Andropogon, Trachypogon, Paspalum). Среди злакового покрова часто доминируют низкорослые деревца Curatella americana, Byrsonima, которые обычны также и в саваннах северной части материка. В понижениях обычны заросли пальм (Mauritia vinifera, Copernicia cerifera, Orbignya). Кампос серрадос имеют весьма большой набор древесных пород, при этом многие глубоко укореняющиеся и вечнозеленые виды отличаются очень интенсивной круглогодичной транспирацией. Следовательно, многие виды этого сообщества могут существовать только за счет богатых резервов влаги в глубоких почвенных горизонтах и не приспособлены к длительной засухе.
В отличие от кампос серрадос вечнозеленые растения области каатинга в засушливой северо-восточной части Бразилии имеют пониженную транспирацию, особенно в период засухи, когда устьица листьев бывают закрытыми. Наиболее сомкнутые древесные сообщества образованы главным образом листопадными породами, среди которых часто встречаются виды со вздутыми стволами, покрытыми колючками. Таковы каваниллезия (Cavanillesia arborea), хоризия (Chorisia), Ceiba pubiflora. Все эти деревья относятся к семейству бомбаксовых, куда включается также баобаб, американским аналогом которого следует, видимо, считать хоризию (барригуда). Кроме того, в лесах этого типа очень заметны различные бобовые из подсемейства мимозовых (Piptadenia macrocarpa. Mimosa) и цезальпиниевых (Caesalpinia pyramidalis).
В нижних ярусах злаки обычно отсутствуют, зато обильны колючие кустарники и суккуленты (кактусовые, молочайные, бромелиевые, толстянковые). Колючие кустарники и суккуленты без древесного яруса распространены более всего, они развиты на маломощных каменистых почвах и выходах коренных пород. Здесь в изобилии встречаются опунции нескольких видов (Opuntia, Tacinga), а также разнообразные цереусовые кактусы: Cereus leucostela, С. jamacaru, Pilosocereus gounelli (часто образуют почти чистые заросли на каменистых обнажениях), Zehntnerella squamulosa, некоторые Melocactus, Stephanocereus leucostele, Micranthocereus polyanthus, Facheiroa ulei, Austrocephalocereus dybowskii. Последние четыре вида кактусовых монотипические и вдобавок эндемичные для области северо-востока Бразилии. Это служит свидетельством того, что каатинга имеет длительное автохтонное развитие.
Некоторое сходство с каатингой имеют и сообщества в области Гран-Чако; органический мир этой территории еще сравнительно мало изучен. Здесь наблюдаются очень большое разнообразие сообществ и богатая флора. В пределах Чако происходит постепенная смена сезонно-засушливых тропических сообществ субтропическими. Разнообразные тропические сезонно-засушливые биоценозы есть также в Центральной Америке, в Вест-Индии и на юге Мексики.
Гетеротрофы первого порядка
Групповой состав гетеротрофов в тропических переменно-влажных сообществах очень сходен с соответствующим набором для вечнозеленых лесов. Однако здесь обычно представлены свои специфические виды или роды. Часть видов, особенно часто среди беспозвоночных животных, оказывается общими с гилеей данного региона. Общее разнообразие фауны падает, хотя в отдельных группах может и возрастать за счет специализированных форм, приспособленных к переживанию неблагоприятного сезона года.
В течение засушливого периода деятельность почвенных микроорганизмов приостанавливается, активность животных-сапрофагов также снижается. В сезонно-сухих лесах обычно хорошо выражен подстилочный горизонт, который дает приют различным потребителям опадающей фитомассы. Как и в гилее, важнейшей группой сапрофагов в дождезеленых лесах и саванновых редколесьях являются термиты. В этих сообществах наличие термитов особенно заметно благодаря внушительным размерам их построек - термитников, высота которых нередко достигает 2-3 м, в исключительных случаях - 9 м а диаметр - 30 м1. Впрочем, чаще всего «фон» образуют сравнительно небольшие башенки, выступающие из-под земли не более чем на 1 м. Таковы, например, остроконические сооружения Amitermes vitiosus и Tumulitermes hastilis в Северной Австралии, придающие своеобразный колорит ландшафту к северу от пустынь (севернее города Алис-Спрингс). В ряде районов обращают на себя внимание термитники грибовидной формы, причем размер «гриба» может колебаться от 10-20 см до 2 м. Часть видов термитов саванн, редколесий и дождезеленых тропических лесов .обходится без надземных башен и ведет сравнительно скрытный образ жизни, проводя большую часть времени в почвенной толще. Архитектура термитников может широко варьировать у видов одного рода и отчасти даже у одного вида в разных условиях. Наибольшее влияние на характер термитных построек имеют количество осадков и соотношение песка и глины в грунте, из которого строятся стенки термитников.
На площади 1 га в сезонно-засушливых тропических ландшафтах насчитывают от 1-2 до 2000 термитников. Обычно их бывают десятки и сотни, при этом плотность крупных колоний с несколькими миллионами особей каждая (представители макротермитин или назутитермитин) обычно менее 10 на 1 га. Надземные постройки занимают от 0,1 до 30% поверхности, чаща - около 0,5-1%. Для саванн Заира к югу от массива дождевых лесов подсчитанная масса субстрата в термитниках доходила до 2400 т/га, что соответствует слою почвы в 20 см на этой площади. По другим данным, общая масса субстрата термитников на 1 га значительно меньше: в Нигерии для Macrotermes bellicosus в саваннах - от 24 до 306 т; близкие цифры отмечены в саванне Южной Африки для Trinervitermes trinervoides. В разных участках саванновых редколесий тропической Австралии наблюдаются следующие сочетания видов по степени участия в общей массе термитников на единицу площади: 1) Tumulitermes hastilis - 30, Drepanotermes rubriceps - 21, Amitermes vitiosus - 10, Nasutitermes triodiae - 1,5; всего 52,5 т/га, или же 2) Nasutitermes triodiae - 3,5, Coptotermes acinaciformes - 1,5, Tumulitermes hastilis - 1, Amitermes vitiosus - 0,4; всего 6,4 т/га. Как видим, картина довольно разнообразная, хотя во всех случаях несомненно значительное влияние на эдафические условия строительной деятельности термитов. Это становится еще более очевидным, если вспомнить, что термиты также энергично сооружают надземные и подземные галереи.
В лесах миомбо из брахистегии на юге Заира общая численность всех термитов, сооружающих надземные постройки, определена в 338 экз./м2. Численность обитающих здесь же других (подземных) термитов равна 500 экз./м2. Общая биомасса всех термитов составляет 11 г/м2. В других местообитаниях этого же района численность только подземных видов достигает 1800 экз./м2. Это составляет обычно около половины от всех крупных почвенных членистоногих. Близкие данные можно найти и в работах по термитам саванн Восточной Африки. Обычно в одном небольшом районе саванн, редколесий или листопадных тропических лесов можно обнаружить 20-30 видов термитов, из которых 10-15 ведут скрытный, подземный образ жизни или обитают в стволах деревьев, а остальные сооружают заметные термитники. Среди всей конкретной фауны термитов относительно обильными оказываются лишь 5-6 видов, на которые приходится около 90% всего населения термитов. Так, на одном участке дождезеленого тропического леса в Нигерии отмечено 19 видов, из них 9 сооружают термитники.
Среди последних абсолютно преобладают лишь три вида: Trinervitermes geminatus, Macrotermes bellicosus, Odontotermes sudanensus. Среди скрытноживущих по обилию и встречаемости резко выделяются два вида: Pseudacanthotermes militaris, Micro-termes sp. В расположенных здесь же осветленных редколесьях и злаковниках абсолютно доминирует в первой группе Т. geminatus, а во второй - Microtermes sp. (тот же вид, что и в лесу) и Anoplotermes sp. (особенно обилен в злаковниках). При этом в редколесьях найдено 17 видов (8 сооружают термитники), а в злаковнике 12 видов (6 сооружают термитники). Сходные соотношения характерны и для других регионов, включая разные материки.
Запасы биомассы, численность, чистая продукция и потребление у населения термитов переменно-засушливых сообществ тропиков весьма сходны с соответствующими величинами для вечнозеленых лесов. Следует, однако, отметить, что в редколесьях и саваннах относительно понижается доля гумусоядных видов. Кроме того, становится заметной группа, потребляющая не только высохшую, но и живую массу травянистого яруса. Таким образом, часть термитного населения включается уже в группу хлорофитофагов, потребителей живой фитомассы.
Большинство термитов дождезеленых лесов и саванн относится к особым видам тех же родов, что представлены в гилее. Это же справедливо и для большинства других сапрофагов, групповой состав которых приведен выше для вечнозеленых тропических лесов. Отмечается лишь общее уменьшение видового разнообразия и появление сравнительно небольшого числа видов-доминантов среди разных групп организмов. В целом же такие группы, как нематоды, клещи-орибатиды, коллемболы, черви-мегасколециды, крупные кивсяки, различные тараканы, сверчки, уховертки весьма обильны и в дождезеленых лесах, и в саванновых редколесьях различных материков.
По сравнению с дождевыми лесами относительно возрастает доля хлорофитофагов, приуроченных к почвенно-подстилочному ярусу (преимущественно ризофаги, т. е. поедающие корни живых растений) и к травянистому ярусу. Характерно, что многие группы почвообитающих беспозвоночных животных (особенно часто личинки различных жуков) переходят от питания детритом к потреблению корней из-за необходимости восполнить водные потери. Благодаря этому один и тот же вид во влажный и в сухой сезоны может переходить из одной трофической группы в другую. Этот сдвиг все более усиливается по мере аридизации климата.
Среди гетеротрофов первого порядка ряд групп становится намного заметнее и обильнее по сравнению с гилеей. Это относится, например, к наземным моллюскам, саранчовым, грызунам и копытным. В Австралии последние заменены крупными травоядными сумчатыми - кенгуру и валлаби. Все перечисленные группы питаются травой или зеленью кустарников, а грызуны - также луковицами и корневищами геофитов. Наличие запаса высококалорийных подземных органов сразу же приводит к появлению роющих грызунов. Обильное плодоношение злаков и других растений с мелкими семенами обусловливает увеличение численности и зерноядных птиц. В первую очередь это касается ткачиковых (семейство Ploceidae) в Африке, Евразии и Австралии, овсянковых в Евразии и Америке (Emberizidae). Среди муравьев в заметном количестве появляются виды, питающиеся семенами трав (муравьи-жнецы), общее обилие этой группы также повышается по мере аридизации климата.
По сравнению с влажнолесными тропическими сообществами наиболее резкое изменение в составе, количестве и ярусном распределении хлорофитофагов можно видеть в биоценозах с хорошо развитым злаковым покровом (т. е. в саваннах, редколесьях и чистых злаковниках). Обилие зеленой массы и семян трав в приземном ярусе создает концентрацию потребителей этих кормов. На всех континентах в первых рядах этих потребителей стоят саранчовые, часто образующие многовидовые группировки. С началом роста злаков большинство саранчовых появляется в виде молодых личинок, а к концу влажного периода и в начале сухого выросшие насекомые откладывают в почву кубышки с яйцами. К концу сухого сезона встречаются взрослые особи, но обычно в небольшом количестве. Кроме того, разные виды сменяют друг друга, образуя сезонные аспекты населения с доминированием то одних, то других группировок. Массовые саранчовые саванн чаще близкородственны пустынным видам, нежели влажнолесным. Очень характерно присутствие немногих представителей, способных к скоплениям и миграциям (тистоцерка, красная и бурая саранча в Африке и др.).
При надземной сухой фитомассе травяного яруса в 0,5-0,9 кг/м2 живая масса саранчовых и других прямокрылых равна обычно 0,5-1 г/м2. Масса всех беспозвоночных-хлорофитофагов травяного яруса, видимо, достигает 1-2 г/м2. Обычно это больше, чем суммарная живая биомасса обитающих здесь же позвоночных-травоядов - грызунов, копытных и т. п. Их суммарная масса в большинстве случаев близка к 0,1-0,5 г/м2.
Заменяющие копытных в саваннах Австралии красные кенгуру (они живут и в полупустынях) обычно имеют биомассу 0,01-0,1 г/м2. Вместе с грызунами, видимо, общая биомасса всех травоядных зверей приблизится к 0,1-0,5 г/м2. Однако в саваннах и редколесьях Восточной и Южной Африки за счет крупных зверей биомасса позвоночных фитофагов может возрастать до фантастических величин в 2-3 г/м2 при том же запасе фитомассы злаков (например, в Национальном парке Вирунга при фитомассе сухих трав в 0,6-0,7 кг/м210 видов копытных и слоны имеют живую зоомассу около 2,4 г/м2).
Пока еще нет удовлетворительно систематизированных данных об относительных запасах биомассы и потоках энергии в различных экосистемах тропических переменно-влажных районов. Но уже сейчас можно констатировать, что относительное участие беспозвоночных и позвоночных животных в общей зоомассе хлорофитофагов может широко варьировать. В Африке при повышении обилия копытных численность грызунов и зайцеобразных падает, и наоборот. Такие же взаимные компенсации, скорее всего, имеют место и между другими животными одной трофической группы. Самые высокие общие запасы обычно достигаются многовидовыми комплексами потребителей живой фитомассы зеленых растений.
Антофилы-опылители и животные-распространители семян принадлежат преимущественно к тем же группам, что и в гилее. Особенно много совпадений можно видеть в лесных сообществах, но даже в саваннах мы находим различных пчелиных (в частности, пчелы-плотники ксилокопы), птиц и рукокрылых-антофилов, плодоядных рукокрылых (в Австралии к ним присоединяются сумчатые мелких размеров). Отметим также большую роль термитников, так как на них (разрушенных или жилых) часто в первую очередь поселяются древесные растения, которые находят здесь наиболее подходящие условия для прорастания семян среди покрова злаков.
Гетеротрофы высших порядков
Ряд паразитов, служащих возбудителями болезней человека и позвоночных животных, весьма обычны в пределах тропических переменно-сухих лесов или саванн. Часть таких возбудителей близка или идентична соответствующим формам из зоны экваториальных лесов (например, малярийные плазмодии и их переносчики - комары из рода Anopheles). Некоторые более свойственны именно сообществам переменно-засушливых тропических районов. Так, например, вероятность заболевания мочеполовым шистозоматозом в Африке наиболее высока в пределах саванн и редколесий вокруг массива влажных экваториальных лесов. Сходная картина наблюдается и для кишечного шистозоматоза. Распространение гельминтозов связано с наличием определенных видов моллюсков (промежуточных хозяев), которые не могут жить в кислой и слабо минерализованной воде гилей и появляются лишь в водоемах саванн и редколесий. Большое количество мухи-цеце (виды из рода глоссина - Glossina) в Африке также приходится на некоторые районы саванн или редколесий (включая леса миомбо). Отсюда - и сходный ареал трипаносом, вызывающих сонную болезнь (гамбийского и родезийского типов). Аналогичные примеры можно было бы привести и по другим материкам.
По сравнению с гилеей общая численность различных насекомых-паразитоидов (наездники, некоторые осы, мухи-тахины и т. п.) увеличивается. Возрастание числа потребителей, злаков в первую очередь и приводит к увеличению особей паразитоидов и хищников. Так, большое число саранчовых вызывает вспышку численности целой группы насекомых, поедающих яйца в кубышках. Обычно таких насекомых также, причисляют к паразитоидам, хотя во многом они стоят уже ближе к хищникам. Таковы, например, личинки мух-жужжал (Bombyliidae) или жуков-нарывников (виды рода Mylabris). Особенно многочисленны жуки-нарывники. Сами жуки питаются растительной пищей. Они хорошо заметны, но более крупные животные не поедают их из-за содержащегося в их теле кантаридина, вызывающего нарывы. Из яиц нарывника выходит мелкая подвижная личинка (триунгулин), которая активно проникает в кубышку саранчовых. После линьки триунгулин превращается в медлительную червеобразную личинку, которая постепенно поедает все содержимое кубышки. Нарывники, как и жужжала, могут также паразитировать и на пчелиных. Жужжала в стадии имаго питаются нектаром, личиночное их развитие весьма сходно с развитием нарывников: личинка первого возраста также подвижна и резко отличается от личинок старших возрастов, которые уже успели проникнуть в кубышку с запасом пищи. Взрослых особей и личинок саранчовых поедает несметное количество врагов - от хищных мух-ктырей (семейство Asilidae), скорпионов, пауков до птиц, ящериц, змей или зверей.
Как и в дождевых лесах, наиболее мощной и всепроникающей группой хищников среди беспозвоночных являются муравьи. Очень многие виды муравьев саванн или сухих тропических лесов имеют близких родственников в гилее. Среди растений и здесь можно найти мирмекофильные виды (например, виды акаций Африки).
Переход многих термитов к обитанию в прочных башнях-термитниках отнюдь не защитил их от специализированных потребителей, о которых уже упоминалось при описании гетеротрофов высших порядков дождевых лесов. Млекопитающие-термитофаги нередко заодно поедают и муравьев, хотя термиты все же составляют большую часть их пищи. Как правило, саванновые виды крупнее своих влажнолесных родственников и ведут наземный, а не древесный образ жизни. В Южной Америке это большой муравьед, несколько видов броненосцев, в Африке и Азии - панголины (ящеры), а в Африке - трубкозуб (Orycteropus afer), в Австралии - сумчатый муравьед.
Гетеротрофы высших порядков решительно преобладают среди амфибий и рептилий, весьма богато представленных в переменно-засушливых зонах всех материков. Обычно эти животные наиболее активны в дождливый сезон. Многие из них засуху переживают совершенно без пищи, зарывшись в почву или спрятавшись в другое убежище. Как и в гилее, среди этих животных есть специализированные термитофилы (амфисбены, слепозмейки, некоторые сцинки), а также сохраняется сравнительно много древесных форм. Некоторые древесные амфибии (квакши, веслоногие и т. п.) в сезон дождей обитают даже среди высоких стеблей злаков.
Обилие копытных, грызунов и других крупных травоядных приводит к обогащению фауны крупных хищных птиц и зверей. Особенно яркий пример этому - Африка с ее богатым набором хищных зверей различных размеров и специализации -от громадных львов до мелких виверровых. Не менее богат также набор падалыциков - гиены, грифы, марабу, шакалы. Впрочем, часть из них может и хищничать. Весьма разнообразен комплекс некрофагов и среди беспозвоночных - это различные падальные мухи из семейств каллифорид (Calliphoridae), capкофагид (Sarcophagidae), мусцид (Muscidae), жуки-мертвоеды (Silphidae), кожееды (Dermestidae). Сюда примыкают и многие виды муравьев. Некоторые падалыцики одновременно потребляют и навоз травоядных животных; к копрофагам относится и много других групп (жуки-навозники, ряд двукрылых).
Общие особенности экосистем
На фоне наиболее сходных абиотических параметров экосистем, в первую очередь своеобразного режима тепла и влаги, формируются разноликие, но в чем-то и похожие биотические сообщества. Созданные средствами разных фаун и флор, на разных материках в близких гигротермических условиях, соответствующие биоценозы достигают очень сходных величин первичной продукции и запаса биомассы. При этом биомасса очень сильно изменяется в одном и том же небольшом районе (лес-злаковник). Изменения продуктивности происходят уже с гораздо меньшей амплитудой, обычно они укладываются в пределы 1-3 кг/м2 сухой биомассы в год чистой продукции (см. карты на с. 63-65).
Особенно поражает богатство фауны и населения крупных зверей, что до сих пор можно видеть в различных национальных парках Африки, многие из которых по размерам территории более 1 млн га. В некоторых парках (например, в Серенгети) в течение многих лет проводятся разнообраз<
Дата добавления: 2019-12-09; просмотров: 1325;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
