Регуляция обмена жиров
Процесс мобилизации резервных триглицеридов или липолизстимулируется рядом гормонов, к числу которых относятся адреналин, норадреналин, глюкагон, b-липотропный гормон гипофиза, соматотропин, АКТГ, МСГ, кортизол, тироксин, тестостерон. Многие из этих гормонов являются активаторами гормончувствительной липазы липоцитов (триацилглицероллипазы). Для оптимального протекания большинства липолитических процессов необходимо присутствие кортизола, соматотропина и гормонов щитовидной железы. Сами по себе эти гормоны не оказывают прямого влияния на липолиз, а действуют как факторы, стимулирующие действие других гормонов.
Важнейшая роль в мобилизации резервных липидов в организме человека принадлежит адреналину (вместе с норадреналином), который выделяется в жировой ткани нервными окончаниями симпатической нервной системы. Вторым источником адреналина является мозговое вещество надпочечников, откуда адреналин доставляется в жировую ткань с током крови. Вероятно, адреналин из мозгового вещества надпочечников играет важную роль в мобилизации триглицеридов жировой ткани в условиях острого эмоционального стресса.
Адреналин взаимодействует со своим рецептором на наружной поверхности мембраны липоцита с образованием гормон-рецепторного комплекса. В ответ на образование гормон-рецепторного комплекса помощью специального механизма происходит активация расположенной на внутренней поверхности наружной клеточной мембраны липоцита аденилатциклазы - фермента, синтезирующего из АТФ циклическую АМФ (цАМФ). Увеличение внутриклеточной концентрации цАМФ активирует фермент протеинкиназу, которая осуществляет активацию триацилглицероллипазы путем ее фосфорилирования, то есть путем ковалентной модификации фермента. Поскольку скорость липолиза лимитируется активностью триацилглицероллипазы, активация фермента приводит к ускорению гидролиза резервных триглицеридов и увеличению выхода высших жирных кислот и глицерола из липоцита в русло крови.
Гормоны глюкагон, b-липотропин, меланоцитстимулирующий гормон, кортикотропин активируют липолиз в жировой ткани, увеличивая концентрацию цАМФ в липоцитах с помощью механизма, сходного с механизмом активации липолиза под действием адреналина. Интересно, что существуют видовые различия в эффективности функционирования этих регуляторных механизмов: так, у птиц глюкагон является мощным стимулятором липолиза, тогда как липолитический эффект глюкагона у человека крайне незначителен.
Соматотропный гормон не оказывает прямого влияния на скорость расщепления триглицеридов в липоцитах, однако соматотропин увеличивает скорость синтеза аденилатциклазы за счет ускорения процесса транскрипции соответствующего гена. Увеличение содержания аденилатциклазы в липоцитах увеличивает эффект воздействия на жировую ткань таких гормонов как адреналин, b-липотропин и др.
Сходным образом оказывает стимулирующее влияние на липолиз и кортизол, поскольку этот гормон увеличивает содержание в липоцитах другого фермента - гормончувствительной липазы. Кортизол выступает в качестве стимулятора транскрипции гена, ответственного за синтез этого фермента. Повышение же содержания триацилглицероллипазы в липоцитах способствует более быстрому и более выраженному ответу клеток на воздействие на них гормонов типа адреналина.
Механизм действия тироксина на жировую ткань не совсем ясен. Известно, что этот гормон способствует более эффективной передаче стимулирующего сигнала с гормон-рецепторного комплекса на аденилатциклазу, в результате чего при воздействии на липоциты гормонов типа адреналина происходит более быстрая активация липолиза в этих клетках.
Основным гормоном, тормозящим липолиз в жировой ткани, является инсулин. Инсулин снижает содержание цАМФ в липоцитах, по-видимому, за счет активации фосфодиэстеразы, переводящей цАМФ в обычную АМФ. Снижение концентрации цАМФ в клетках приводит как к инактивации протеинкиназы, так и к активации фосфопротеинфосфатазы, в результате чего происходит дефосфорилирование гормончувствительной липазы с ее инактивацией и последующим торможением липолиза.
Простагландины также снижают содержание цАМФ в липоцитах с последующим торможением липолиза в клетках.
В период абсорбции в клетках различных органов и тканей активно идет липогенез. Во внутреннюю среду организма из кишечника поступают глюкоза и другие моносахариды, а также триацилглицерины. Моносахариды, поступающие в липоциты или в гепатоциты, используются в ходе липогенеза, являясь как источниками ацетил-КоА для синтеза высших жирных кислот, так и источниками фосфотриоз, необходимых для образования 3-фосфоглицерола. Триглицериды также являются источниками высших жирных кислот и глицерола, поступающих в клетки и в дальнейшем используемыми в качестве субстратов для липогенеза.
Гормоном, стимулирующим липогенез, является инсулин. Он ускоряет поступление глюкозы в клетки и стимулирует ее фосфорилирование, запуская тем самым процесс утилизации глюкозы в клетках. Причем стимулируется как процесс аэробного окисления глюкозы до СО2 и Н2О, так и работа пентозного цикла окисления глюкозы, обеспечивающего клетки восстановительными эквивалентами в виде НАДФН+Н+.
Инсулин активирует работу пируватдегидрогеназного комплекса, что приводит к увеличению образования ацетил-КоА - исходного субстрата для синтеза высших жирных кислот. Инсулин повышает активность фермента ацетил-КоА-карбоксилазы, катализирующего превращение ацетил-КоА в малонил-КоА, также необходимого для синтеза высших жирных кислот. Ускорение окислительного распада глюкозы в клетке приводит также к увеличению в ней концентрации фосфотриоз - 3-фосфоглицеринового альдегида и фосфодигидроксиацетона, используемых для образования 3-фосфоглицерола.
Таким образом, воздействие инсулина на клетки приводит к наработке в них исходных соединений для синтеза триглицеридов. Кроме того, инсулин активирует в клетках глицеролфосфат-ацилтрансферазу - фермент, катализирующий перенос ацильного остатка с КоА на 3-фосфоглицерол первую реакцию метаболического пути синтеза триацилглицеринов. Роль инсулина в депонировании жира показана на рис. 17.
Регуляция активности пируватдегидрогеназного комплекса, ацетил-КоА-карбоксилазы и глицеролфосфат-ацилтрансферазы осуществляется путем координированного процесса ковалентной модификации этих ферментов (фосфорилирование - дефосфорилирование).
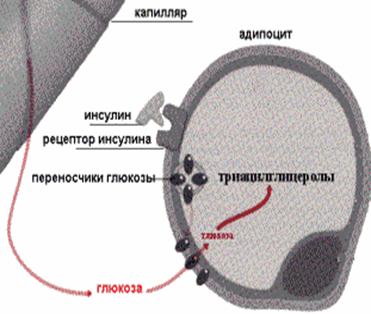
Рис.17. Роль инсулина в депонировании жира
В целом, воздействие инсулина на липоциты приводит, во-первых, к торможению липолиза в клетках, а, во-вторых, к активации в них процесса липогенеза, способствуя тем самым накоплению энергетических резервов в организме в виде триацилглицеринов.
В обмене жиров также большая роль принадлежит нервной системе. При повреждении некоторых ядер гипоталамуса жировой обмен нарушается и происходит ожирение организма или его истощение. Нервная регуляция жирового обмена осуществляется путем прямых воздействий на ткани (трофическая иннервация) или через железы внутренней секреции.
Дата добавления: 2016-06-09; просмотров: 4085;
Поиск по сайту
Узнать еще
- IV. Ликвидация гиповолемии и регуляция водно-солевого обмена
- Vi. регуляция водно-солевого обмена
- Vii. регуляция обмена ионов кальция и фосфатов
- Абсцесс, флегмона подкожножировой клетчатки переднего отдела подподъязычной части шеи
- АВТОНОМНАЯ САМОРЕГУЛЯЦИЯ ФЕРМЕНТАТИВНЫХ ПРОЦЕССОВ
- Алгоритм энергообмена в колебательных системах
- Аллостерическая регуляция каталитической активности ферментов
- Анаэробный гликолиз: понятие, этапы, общая схема, последовательность реакций, регуляция, энергетический баланс. Регуляция гликолиза

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
