Глава 8. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА И МОРФОФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ МЫШЛЕНИЯ ЖИВОТНЫХ.
Сравнительная характеристика уровня развития элементарного мышления у животных разных таксономических групп. Некоторые показатели уровня развития мозга (относительные размеры его высших отделов, а также сложность нейронного строения и межнейронных связей), которые коррелируют со степенью развития элементарной рассудочной деятельности. Данные о дифференцированном влиянии разрушения одних и тех же структур на рассудочную деятельность и способность к обучению. Показано, что животным, обладающим хорошо развитым мозгом, доступны более сложные по логической структуре и более разнообразные задачи. Характеристики рассудочной деятельности наиболее продвинутых животных, степень их сходства с психикой человека и отличия от психики других животных. Обсуждение параллелизма в развитии рассудочной деятельности представителей разных классов.
Современное исследование любого аспекта поведения только тогда может считаться полноценным, если авторы рассматривают его развитие, механизмы, эволюцию и адаптивное значение, не ограничиваясь наблюдениями и описаниями. В предыдущих главах был рассмотрен практически весь диапазон форм элементарного мышления, доступных животным. Одни формы мышления достаточно примитивны и встречаются у низкоорганизованных животных, другие — более сложные и доступны видам, наиболее продвинутым в эволюционном развитии. Интересно рассмотреть, на каких этапах филогенеза они возникли, как происходило их усложнение в эволюционном ряду, а также выяснить, какие особенности строения мозга обеспечивают проявление того или иного уровня развития рассудочной деятельности.
8.1. «Сложное обучение» и уровень развития животных.
В многочисленных исследованиях было неоднократно продемонстрировано, что скорость, прочность и «точность» формирования обычных УР (как классических, так и инструментальных) достаточно близки у позвоночных самого разного филогенетического уровня и практически не зависят от сложности строения мозга. Чтобы оценить степень развития более сложных когнитивных функций, необходимо было разработать другие процедуры обучения животных, которые требовали бы улавливания общего «правила», «алгоритма» решения серии однотипных задач. Предполагалось, что их использование могло бы выявить различия в обучении и когнитивных процессах, которые коррелировали бы со сложностью строения мозга.
В лаборатории Л. Г. Воронина в 60-е годы на животных разных таксономических групп (рыбах, пресмыкающихся, птицах и млекопитающих) исследовали скорость формирования «цепных» УР. Оказалось, что «цепи» УР легче вырабатываются и дольше сохраняются у животных с более развитым мозгом.
Методом многократных переделок УРуже в первых работах убедительно показано, что успешность обучения в целом зависит от уровня организации мозга животного, по крайней мере в пределах крупных таксономических групп (Биттерман, 1973; см. 3.3.1). При последовательных переделках сигнального значения стимула формирование каждого следующего УР у многих млекопитающих (в том числе у крыс) и некоторых видов птиц ускоряется, т.е. число ошибок при каждом новом сигнальном значении стимулов постепенно и достаточно заметно снижается. В отличие от них у рыб такого ускорения практически не происходит. Таким образом, способность уловить общее правило, лежащее в основе теста, проявляется у некоторых животных с относительно примитивным мозгом (крысы, голуби).
Формирование установки на обучение(см. 3.3.3) также позволило выявить различия в способности к обучению у животных разных видов, коррелирующие с уровнем развития мозга (рис. 8.1).
Как видно из рисунка, у большинства видов приматов установка на обучение формируется после выработки 150—200 дифференциро-вок. Иными словами, в этот период доля правильных выборов уже при втором предъявлении новых стимулов (т.е. без дополнительного обучения) доходит до 90%. Несколько больше таких серий обучения требуется беличьим саймири, еще больше — мармозеткам и кошкам. В отличие от них у крыс, кур, голубей и белок правильные ответы при втором предъявлении новой пары стимулов превышали случайный уровень не более чем на 10—15% даже после выработки 1500 различных дифференцировок. Для крыс этот тест оказался менее доступным, чем многократная переделка УР.
Врановые птицы — американские сойки (Суапосilla cristata) и вороны (Corvus brachyrhynchos), а также скворцы-майны (Gracula religiosa) по скорости формирования установки на обучение превосходят кошек и саймири и приближаются к мартышковым обезьянам. Для них характерны также сходные стратегии решения теста (см. ниже).
Скорость формирования установки на обучение соответствует уровню организации мозга: низкая — у грызунов, выше — у хищных млекопитающих и очень высокая — у приматов в целом.
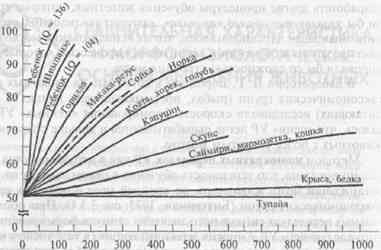
Рис. 8.1. Сравнительная характеристика способности животных разных видов к формированию установки на обучение.
По оси ординат — доля (%) правильных выборов при второй пробе, по оси абцисс — предъявление задачи. По Hodos, Campbell, 1979, с дополнениями.
Вместе с тем внутри отряда приматов существуют определенные различия по этому показателю. Наиболее успешно формируют «установку» человекообразные обезьяны, причем шимпанзе опережают в этом отношении других антропоидов, включая горилл, и даже детей с низким коэффициентом интеллектуального развития. Это служит одной из важных иллюстраций широко распространенного представления об исключительной близости шимпанзе к человеку (Rumbaugh et al., 2000).
Наряду со столь очевидными различиями показателей решения, которые обнаружены у шимпанзе и грызунов, во многих случаях животные с заведомо по-разному организованным мозгом (например, кошки и макаки) демонстрируют сходные количественные показатели формирования установки. Однако различия между ними выявляются четко, если обратиться к «качественному» анализу, т.е. к сопоставлению стратегий в решении этого теста. Такой анализ провел Д. Уоррен (Warren, 1977). Кошки действовали достаточно примитивно. Если при первом предъявлении новой пары стимулов кошка случайно выбрала «правильный» стимул, то дальше действовала безошибочно, т.е. придерживалась стратегии «преуспел — делай так же» (win-stay). При неудачном выборе кошка не могла адекватно использовать полученную информацию и в следующий раз выбирала наугад, не используя стратегию «проиграл — измени тактику» (lose-shift), а дальше обучалась по принципу «проб и ошибок».
В таких же опытах макаки-резусы вели себя иначе. Даже если при первом предъявлении новой пары стимулов их выбор был неудачным, то при следующих пробах они практически всегда действовали правильно. Иными словами, «неуспех» воспринимался макаками не как ошибка, после которой надо снова пробовать, а как основание для смены стратегии выбора, т.е. они далее действовали по правилу: «проиграл — измени тактику». К тому же, в отличие от кошек, макаки могли гибко переходить от одного правила к другому. Это означало, по-видимому, что они способны полностью уловить принцип задачи. Такую же более совершенную стратегию формирования «установки» проявили представители врановых птиц — сойки: они реагировали на новые стимулы правильно независимо от того, был ли выбор в первой пробе положительным или отрицательным (Kamil, 1988).
Межвидовые различия обнаруживаются и в скорости обучения «выбору по образцу». Если крысам и голубям требуются сотни сочетаний для формирования и упрочения реакции выбора «по сходству с образцом», то человекообразным обезьянам нужен гораздо меньший период тренировки, а в некоторых случаях им достаточно первых проб. Методика обучения навыку выбора «по отличию от образца» широко используется в исследованиях обучения ипамяти у грызунов; она оказалась пригодной и для оценки особенностей когнитивных процессов у трансгенных мышей (см. также 9.5).
При сравнении способностей животных разных видов к обучению простым условным рефлексам различий не обнаруживается. В тестах на способность к сложным формам обучения, когда животное должно уловить (понять) общее правило выбора, большего успеха добиваются животные с более высокоорганизованным мозгом.
8.2. Сравнительная характеристика уровня элементарной рассудочной деятельности (элементарного мышления) у животных разных таксономических групп.
К началу 70-х годов XX в. сформировались экспериментальные подходы, позволившие проводить систематические сравнительные исследования рассудочной деятельности животных. Их особенностью было использование животных большого числа видов в стандартных условиях с применением единых, стандартных тестов, результаты которых доступны точной количественной оценке. Это позволяло сопоставлять показатели решения теста животными разных таксономических групп и анализировать специфику механизмов (стратегии), лежащих в основе решения такой задачи, подобно тому как это было сделано при сопоставлении стратегий решения теста на «установку» (см. 8.1).
Способность к экстраполяции.Наиболее полная сравнительная характеристика рассудочной деятельности была получена с помощью теста на способность к экстраполяции, а также некоторых других элементарных логических задач, разработанных. Л.В. Крушинским (1986). Задача межвидовых сравнений облегчалась тем, что существовали методы точной количественной оценки результатов этих тестов (см. гл. 4).
В главе 4 (см. 4.6.2; рис. 4.12) были описаны основные результаты таких опытов, проведенных на представителях всех классов позвоночных: рыбах (4 вида), земноводных (3 вида), пресмыкающихся (5 видов), на 15 видах млекопитающих и 13 видах птиц (см.: Крушинский, 1986). (Подробное изложение этих данных можно найти в работах Крушинского (1986), Зориной (1997), Очинской (1971), Полетаевой (1998), Флесса и др. (1974; 1987).
Как показывает рис. 4.12А, способность к экстраполяции имеется у многих животных. Совершенно не способными к решению этой задачи оказались только рыбы и земноводные. По данным Е. И. Очинской (1971), задачу на экстраполяцию успешно решали пресмыкающиеся — черепахи, кайманы и зеленые ящерицы. Отметим, что у черепах была выявлена также способность к обобщению отвлеченных пространственных признаков (см. 5.5.3).
Наличие у пресмыкающихся способности к экстраполяции и обобщению свидетельствует, что зачатки этих форм элементарного мышления сформировались на относительно ранних этапах филогенеза.
Наиболее полно способность к экстраполяции была охарактеризована у млекопитающих. В пределах этого класса можно наблюдать закономерное улучшение решения большинства тестов на рассудочную деятельность (см. ниже). Так, у грызунов в целом способность к экстраполяции крайне ограниченна, хищные млекопитающие прекрасно экстраполируют, у приматов эта способность не оценивалась, а у дельфинов она высоко развита.
Тесты на оперирование эмпирической размерностью фигур и Ревеша—Крушинского.Согласно взглядам Л. В. Крушинского, способность к экстраполяции направления движения пищевого или другого биологически значимого раздражителя отражает лишь одну из возможных сторон рассудочной деятельности животных. Другой тест — оперирование эмпирической размерностью фигур (см. 4.6.3) — основан на понимании геометрических свойств предметов. Его использование позволило углубить сравнительную характеристику рассудочной деятельности исследованных видов животных. Его могут решить животные лишь немногих видов (см. 4.6.3.1). Удивительно, что с ним не справляются хищные млекопитающие (за исключением медведей). Врановые птицы решали задачу на уровне, близком к обезьянам, медведям и дельфинам. Эти опыты, так же как и результаты исследования способности врановых птиц к обобщению и символизации, свидетельствуют о сходстве уровня рассудочной деятельности этих птиц и приматов.
Тест Ревеша—Крушинского был предназначен для выявления способности животных экстренно определять положение спрятанной приманки на основе информации о ее перемещении, полученной в ходе теста (см. 4.7). Все исследованные виды (крысы, врановые птицы, некоторые виды низших обезьян и человекообразные обезьяны) ведут себя практически одинаково — безошибочно решают задачу лишь в единичных случаях, однако все животные (и крысы, и приматы) способны оптимизировать поиск в пределах первого же теста.
Наряду со способностью к экстренной реорганизации независимых навыков (см. 4.8.1) тест Ревеша—Крушинского — это еще одна форма рассудочной деятельности, доступная низкоорганизованным животным — крысам.
«Градации» элементарного мышления.Способность к экстраполяции направления движения пищевого раздражителя, исчезающего из поля зрения, обнаружена у представителей пресмыкающихся, млекопитающих и птиц, но выражена в разной степени. На этом основании Л. В. Крушинский выделил несколько градаций в степени ее развития: они различались не только по количественным показателям (от 65% у некоторых линий мышей до 90% у хищных млекопитающих), но и по способности решать различные усложненные варианты этой задачи. Появление данных о способности к решению задачи на оперирование эмпирической размерностью фигур позволило дать еще более подробную характеристику уровня развития элементарного мышления (см. ниже).
Л. В. Крушинский высказал гипотезу, что усложнение рассудочной деятельности животных в процессе эволюции происходило за счет увеличения числа «эмпирических законов», которыми могут оперировать животные, и, следовательно, росло число элементарных логических задач, которые они способны решать.
Исходя из этого, Крушинский полагал, что для сравнительной характеристики рассудочной деятельности животных необходимо использовать батареи разнообразных тестов.
Результаты исследования элементарного мышления животных, накопленные к настоящему времени, показали плодотворность и информативность такого подхода.
В главе 4 уже говорилось, что элементарное мышление животных исследуют с помощью двух групп тестов.
Первая из них оценивает способность животного к решению задачи в экстренно сложившейся обстановке, основанному на понимании логической структуры задачи (к таким задачам относится и тест на экстраполяцию). Л. В. Крушинский предложил набор (или батарею) тестов разной сложности для комплексной оценки элементарной рассудочной деятельности животных. Его работы позволили выявить градации таких способностей в ряду позвоночных.
Вторая группа тестов анализирует способности животных к обобщению и абстрагированию. Данные, полученные в экспериментах по обучению многократным переделкам дифференцировочных УР и «установке на обучение», также выявили градации этих способностей у животных разного уровня организации и показали сходный характер различий между разными таксономическими группами.
Млекопитающие.Грызуны характеризуются низшей градацией элементарного мышления. Способность к экстраполяции обнаружена у диких крыс-пасюков (Крушинский и др., 1975), некоторых генетических групп мышей (Полетаева, 1998; см. 9.2 и рис. 9.1) и бобров (Крушинская и др., 1980), причем в большинстве случаев правильные решения лишь незначительно превышают случайный уровень. Тем не менее эти решения по своему механизму принципиально отличаются от обучения сходной задаче (см. ниже) и представляют собой проявления более сложной, чем обучение, когнитивной способности. Наряду со слабой способностью к экстраполяции, у грызунов крайне ограничена способность к обобщению (см. гл. 5), и они не могут формировать установку на обучение. В то же время некоторые когнитивные задачи им доступны — крысы способны к решению задач на экстренную реорганизацию независимо сформированных навыков (см. 4.8) и к оптимизации стратегии при поиске приманки в тесте Ревеша—Крушинского (см. 4.7).
Следующая градация обнаружена у хищных млекопитающих. Все исследованные виды этого отряда (кошки, собаки, волки, лисы, песцы, медведи) успешно решают задачу на экстраполяцию. Это совпадает с их выраженной способностью к формированию установки на обучение (см. выше) и к достаточно высокому уровню обобщений (см. гл. 5). Вместе с тем важно подчеркнуть, что большинство хищных млекопитающих не способны к решению теста на оперирование размерностью фигур. Это объективно отражает специфику их когнитивных способностей и отличие уровня развития хищных от приматов.
Следующую градацию элементарного мышления можно обнаружить у более высокоорганизованных млекопитающих — обезьян и дельфинов (Флесс и др., 1987). Дельфины хорошо экстраполируют направление движения раздражителя, что согласуется с их способностью к быстрому формированию установки на обучение, к высоким степеням обобщения и другим сложным когнитивным функциям (см. также 8.5).
Птицы.В пределах класса птиц обнаружены сходные с млекопитающими градации способности к экстраполяции — от полного ее отсутствия у голубей до высокого ее развития (на уровне хищных млекопитающих и дельфинов) у врановых птиц. Хищные птицы (Falco tinunculus, F. vespertilus, Pernis aviporus и др. виды) занимают промежуточное положение: у них уровень успешных решений при первом предъявлении лишь незначительно (хотя и достоверно) превышает случайный.
Эта характеристика становится более полной и убедительной в сопоставлении с данными по другим видам элементарного мышления у врановых и голубей.
Врановые птицы достигают уровня развития приматов по следующим видам когнитивных тестов:
♦ по скорости и стратегии образования установки на обучение;
♦ по способности к оперированию эмпирической размерностью фигур
(см. 4.6.2);
♦ по возможности образования довербальных понятий (см. 5);
♦ по способности к употреблению символов (см. гл. 5 и 6).
В отличие от них голуби — значительно более примитивно организованные представители класса птиц. Они не способны к решению элементарных логических задач, к формированию установки на обучение и обладают крайне ограниченной способностью к допонятийному уровню обобщения. Тем не менее даже у них проявляется способность к решению наиболее простой задачи—к экстренной интеграции независимо образованных навыков (см. 4.8.1).
Способность к экстраполяции представляет собой относительно универсальную когнитивную функцию, в той или иной степени доступную широкому диапазону видов позвоночных, начиная с рептилий. Таким образом, самые первые и примитивные биологические предпосылки мышления человека возникли на ранних этапах филогенеза позвоночных.
Более высокоорганизованные животные способны к решению большего числа когнитивных тестов и справляются с более сложными логическими задачами. Это соответствует представлениям Л. В. Крушинского о том, что эволюция элементарной рассудочной деятельности животных шла, по-видимому, в направлении увеличения числа «законов», которыми животное способно оперировать.
8.3. Рассудочная деятельность и сложность строения мозга.
Л.В. Крушинский и его сотрудники в 70-е годы XX века предприняли исследование морфофизиологических основ элементарной рассудочной деятельности животных. Эти работы включали сопоставление сложности строения высших ассоциативных структур переднего мозга птиц и млекопитающих с уровнем развития у них способности к рассудочной деятельности. На основе таких сопоставлений была изучена роль отдельных образований мозга в способности к экстраполяции и в способности к обучению. Ряд работ был специально посвящен сравнению способности животных к решению элементарных логических задач и к обучению.
8.3.1. «Цефализация», сложность нейронного строения и уровень рассудочной деятельности.
В лаборатории Л. В. Крушинского изучалась связь сложности организации мозга и общего уровня эволюционного развития в классах птиц и млекопитающих. Нейроморфологические данные, накопленные к 70-м годам XX в., свидетельствовали, что индекс цефализа-ции — относительный объем высших отделов мозга (новой коры у млекопитающих и гипер- и неостриатума у птиц) растет по мере повышения уровня эволюционного развития вида. Л. В. Крушинский (1986) показал, что как у птиц, так и у млекопитающих степень цефализации в пределах каждого класса позвоночных представляет собой существенный параметр, определяющий уровень развития рассудочной деятельности.
А. Портман (Portmann, I946) получил следующие величины индексов относительного объема полушарий птиц: голубь — 4,0; курица — 3,27; утка (Anas platirhinchos) — 6,08; сокол (Falco tinninculus) — 8,24; канюк (Buteo buteo) — 9,78; сорока (Pica pica) — 15,81; грач (Corvus frugilegus) — 15,68; ворона (ы) — 15,38.
У млекопитающих также обнаруживается зависимость между уровнем развития элементарной рассудочной деятельности и относительным размером мозга. Л. В. Крушинский приводит следующие величины квадратического показателя головного мозга (по Я. Я. Рогинскому) для ряда видов млекопитающих: мышь — 0,0088; крыса — 0,0123; кролик — 0,0705; кошка — 0,195; собака — 0,464. Приматы и дельфины обладают наиболее дифференцированным и крупным мозгом среди млекопитающих.
В ряду млекопитающих происходит увеличение площади ассоциативных зон коры больших полушарий, в частности префронтальной (лобной) области. Это тоже является индикатором усложнения строения высших отделов мозга. Такая же закономерность описана и в отношении ассоциативных областей мозга птиц.
В этой связи следует отметить и еще одно немаловажное обстоятельство. Сравнительные исследования Л. В. Крушинского и его сотрудников (1986) показали, что нет прямой и непременной связи между степенью развития элементарной рассудочной деятельности и наличием новой коры. Мозг птиц построен по иному плану, чем мозг млекопитающих. В процессе филогенеза особого развития у них достигли новые, отсутствующие у млекопитающих, отделы стриатума (гипер- и неостриатум— wulst), в то время как у млекопитающих сформировалась новая кора. Именно за счет этих отделов стриатума увеличился объем полушарий и переднего мозга врановых птиц
Наряду с увеличением относительных размеров высших отделов мозга, другим важным фактором, определяющим степень развития рассудочной деятельности, следует считать сложность нейронной организации мозга. В филогенетическом ряду позвоночных наблюдается увеличение разнообразия нейронного строения мозга, с прогрессирующим увеличением числа так называемых звездчатых нейронов (Богословская, Поляков, 1981), а также усложнение систем контактов между нейронами. Были выявлены особенности цитоархитектони-ки переднего мозга вороны и голубя (Крушинский и др., 1985; Воронов, 1996).
Особенно сложным строением отличаются нейроны гиперстриа-тума вороны. Они имеют более длинные и более извилистые дендри-ты, что определяет большее число контактов с другими клетками. Количество шипиков на дендритах также значительно больше, чем на дендритах нейронов мозга голубя.
Специфическая особенность строения мозга птиц — так называмые мультинейронные комплексы (Богословская, Поляков, 1981). Это клеточные ассоциации сложной пространственной структуры, состоящие, по-видимому, из функционально связанных нервных элементов. Исследования Д. К. Обухова (Обухов, 1996; Андреева, Обухов, 1999) демонстрируют конкретное строение таких мультинейронных комплексов: у ворон они могут включать до 20 нейронов, у голубей до 10.
В двух классах позвоночных — млекопитающих и птиц — усложнение строения мозга в ряду видов коррелирует с повышением уровня развития их элементарного мышления. Это прослеживается и в увеличении числа тестов, которые более развитые животные способны решать, и в повышении уровня сложности этих тестов.
8.3.2. Повреждение участков мозга и способность к экстраполяции.
Анализ морфофизиологических основ рассудочной деятельности проводился в 60—70-е годы в лаборатории Л. В. Крушинского. В опытах Д. А. Флесса (1974) был начат фармакологический анализ способности к экстраполяции. Было показано, что введением холинолитиков она блокировалась, но сохранялась при использовании препаратов, выключающих катехоламинергическую нейромедиаторную систему мозга (даже при использовании очень высоких дозировок).
Более подробно была исследована роль различных структур мозга у животных, обладающих разной способностью к экстраполяции, в опытах с их повреждением (удалением, экстирпацией). Для анализа последствий разрушений участков мозга были выбраны представители разных классов, в пределах класса — разные виды, в пределах одного вида — разные группы животных. Часть экспериментов проводили на животных, взятых в опыт впервые, а часть — на животных, имевших до операции опыт решения задачи. Разрушались структуры, ответственные за высшие ассоциативные функции и имеющие тенденцию к развитию иусложнению в пределах своего класса: префронтальная кора млекопитающих, комплекс ядер переднего мозга птиц — Wulst и дорсовент-ральная область коры — у пресмыкающихся, поскольку степень их развития коррелирует с уровнем способности к экстраполяции. В «контрольных» опытах был проанализирован эффект равных по объему разрушений других структур переднего мозга, выполняющих более элементарные функции. Эксперименты проводились на собаках (успешно решающих тест на экстраполяцию), кошках (чьи показатели были ниже), воронах (имеющих высокие показатели во всех тестах), курах (тест не решают), болотных черепахах Emis orbicularis, успешно экстраполирующих (Зорина, Попова, 1978; Зорина, Федотова, 1981; Адрианов, Молодкина и др., 1987; Очинская и др., 1988).
У млекопитающих, решавших задачу впервые,способность к экстраполяции нарушалась при повреждении префронтальной коры (высшей ассоциативной зоны) и не страдала при повреждении других областей коры.
У врановых птиц, которым тест был также предъявлен впервые, аналогичные нарушения наблюдались при повреждении Wulst (комплекс ядер гиперстриатума (Karten, 1991), который считают функциональным аналогом префронтальной коры млекопитающих), и отсутствовали при контрольном разрушении гиппокампа (старой коры).
У рептилий (черепах) способность к экстраполяции нарушалась при экстирпации дорсовентральной области коры (зоны перекрытия сенсорных проекций разных модальностей). В мозге рептилий она выполняет как ассоциативные, а так и проекционные функции. При разрушении чисто проекционной области выполнение теста не нарушалось.
После разрушений участков мозга, имеющих высшие ассоциативные функции, все оперированные животные первое время решали задачу на экстраполяцию чисто случайно. Однако по мере его многократных повторений доля правильных обходов ширмы постепенно увеличивалась, т.е. животное обучалось навыку обхода ширмы со стороны движения корма (см. также ниже).
Если разрушение производили у животных, успешно решавших задачу до операции, способность к экстраполяции в целом сохранялась, хотя доля правильных решений снижалась. Следовательно, разрушение указанных структур не нарушает способности животных к обучению, но практически исключает проявление способности к экстраполяции при первом предъявлении задачи. Сходство эффектов разрушений у представителей всех трех классов отражает, по-видимому, общность процессов, лежащих в основе решения этого теста.
В опытах с птицами в качестве «контрольной» структуры, разрушение которой не должно было изменить уровня решения задачи на экстраполяцию, была выбрана старая кора (гиппокамп). В главе 3 (см. 3.4.3) было показано, что повреждение этой структуры у птиц и млекопитающих существенно ухудшает процесс запоминания. В наших опытах при разрушении гиппокампа поведение ворон (не имевших опыта решения задачи до операции) не изменялось ни в первом, ни при повторных предъявлениях теста на экстраполяцию. У кур (низкий уровень рассудочной деятельности) свойственное им постепенное формирование условно-рефлекторного навыка обхода ширмы после разрушения гиппокампа замедлялось, в то время как при разрушении гиперстриатума их поведение не изменялось.
Сходным образом разрушение хвостатого ядра не ухудшало решения задачи на экстраполяцию у кошек, но ослабило эту способность у крыс (выраженную слабо даже в норме).
У представителей млекопитающих, птиц и рептилий выявлены структуры мозга, в наибольшей степени связанные с осуществлением решения задачи на экстраполяцию. Их разрушение практически исключает проявление способности к экстраполяции при первом предъявлении задачи, ухудшает показатели животных, решавших задачу до операции, но не препятствует обучению при многократных предъявлениях. Различие влияний на рассудочную деятельность и на обучение сходным задачам позволяет говорить об участии в этих процессах разных нейрофизиологических механизмов. Сходство эффектов разрушений у представителей всех трех классов отражает общность процессов, лежащих в основе решения этого теста.
Сопоставление решения задачи на экстраполяцию и обучения сходному навыку проводилось и в других экспериментах.
8.4. Сопоставление способности к экстраполяции и к обучению. Роль экологических факторов в успешности решения тестов.
Решение элементарных логических задач, даже при их повторных предъявлениях, представляет собой самостоятельный феномен, по своей природе отличный от других форм индивидуально-приспособительного поведения, в частности от инструментального обучения. Об этом свидетельствуют отличия в поведении животных в контрольных тестах, которые по своей структуре сходны с элементарной логической задачей, за исключением того, что в них отсутствует логическая структура. «Решить» такой тест при первом предъявлении невозможно, безошибочного выполнения его можно достичь только путем обучения методом проб и ошибок. Выше (см. 4.6.3, рис. 4.16) был описан такой контрольный опыт для теста на оперирование эмпирической размерностью фигур. Подобному контролю подвергали и тест на экстраполяцию.
В поведении мышей при обучении навыку отыскания исчезнувшего корма и при решении теста на экстраполяцию также выявлены существенные различия (Крушинский и др., 1982). Была обнаружена группа мышей, носителей хромосомной мутации (подробнее см. 9.2 и рис. 9.1), у которых доля правильных решений теста на экстраполяцию достоверно превышала случайный уровень. Их поведение сравнивали с мышами линии СВА, которые решали тест на 50%-м случайном уровне.
В тесте на экстраполяцию мышь может следить за направлением исчезновения корма, затем она идет либо в «правильном» направлении — в сторону исчезновения корма, к соответствующему боковому отверстию в стенке камеры, либо в противоположном направлении. Контрольный тест на обучение навыку обходить ширму с определенной стороны проводился в той же камере, что и исследование способности к экстраполяции (см. рис. 4.12); он был сходен с тестом на экстраполяцию по структуре. Мышь также начинала пить молоко из поилки, которую от нее закрывали (т.е. как и в основном тесте прекращали доступ к корму), однако никакого движения кормушки в поле зрения мыши не происходило. Подкрепление мышь находила всегда около одного из боковых отверстий. После этого она снова подходила к центральному отверстию и т.д., 10 раз за опытный день.
Уже после трех дней обучения у мышей обеих групп время подхода к поилке сократилось, у них сформировался инструментальный пи-щедобывательный УР на место подкрепления. В тестах на экстраполяцию за такой же период время подхода к поилке у мышей разных групп изменилось по-разному: у «способных» к экстраполяции мышей с хромосомной мутацией оно практически не изменилось, тогда как у неспособных мышей СВА оно сократилось.
Мыши СВА обучились быстрее перебегать к боковым отверстиям, хотя по-прежнему (и к третьему опыту) они не улавливали логической структуры задачи (доля правильных решений у них осталась на случайном уровне).
Мыши с мутацией (успешно усвоившие такой инструментальный навык в специальном тесте на обучение) к третьему опытному дню в тесте на экстраполяцию не стали бегать быстрее. По-видимому, каждое решение задачи у них осуществлялось не на основе выполнения выученного двигательного навыка, а на основе улавливания логической структуры теста. Такое поведение не обязательно влечет за собой ускорение подхода к пище при повторениях.
Адаптивные реакции поиска пищи у лабораторных мышей могут происходить как на основе способности к экстраполяции, так и на основе более простого механизма, на основе инструментального обучения. Эксперимент позволяет четко разделить эти два механизма.
Прямые свидетельства различий в механизмах обучения и способности к решению элементарных логических задач были получены также в опытах с врановыми птицами. Оказалось, что молодые птицы неспособны к решению задачи на экстраполяцию и ОЭРФ, и такая способность обнаруживается у них не ранее, чем в годовалом возрасте. В то же время способность к обучению — выработке простых и диффе-ренцировочных УР, их сложных «систем» и т.п. — обнаруживается уже у 3-месячных птенцов. Сроки «созревания» способности к элементарному мышлению у ворон совпадают с окончанием процесса миели-низации проводящих путей в переднем мозге и появлением наиболее крупных мультинейронных комплексов (Зорина, Крушинский, 1987; Воронов, 1996).
Особенности экологии вида и способность к элементарному мышлению.При анализе механизмов решения элементарных логических задач неизменно возникал вопрос о том, в какой мере их можно отнести за счет наличия или отсутствия способности к рассудочной деятельности, а в какой они являются результатом каких-то иных механизмов, в том числе частных экологических адаптации. Вопрос чаще всего возникал в связи с анализом решения задачи на экстраполяцию и требовал специального анализа. Данные по способности к экстраполяции у представителей разных систематических групп были сопоставлены с особенностями кормовой специализации.
Рассмотрим некоторые примеры. Одним из аргументов в пользу «экологической» трактовки данных о способности вида к экстраполяции было существование специализации в добыче пищи. В репертуар видоспецифического поведения ряда животных (особенно хищных) входит преследование движущейся добычи. Однако оказалось, что наличие или отсутствие хищнического поведения не является решающим фактором в определении уровня рассудочной деятельности животного. У рыб способности к экстраполяции не обнаружено, хотя тестированы были как хищные виды (цихлидовые), так и травоядные (караси). Напротив, столь же контрастные по способу питания черепахи — Emis orbicularis и Clemmis caspica — хищные виды и Testudo horsfield — травоядный — задачу на экстраполяцию решают успешно (Очинская, 1971).
Отметим, что у целого ряда видов существуют видоспецифичес-кие механизмы, позволяющие им экстраполировать направление (видимого) движения объектов. Такой механизм обеспечивает добычу пищи у амфибий, которые ловят насекомых на лету, однако не помогает им экстраполировать направление движения скрывшегося корма, так как для решения этой задачи необходимо уловить связь между пищевым стимулом и изменением его положения на видимом отрезке движения, а затем принять решение о направлении его движения на скрытом от глаз участке.
Наконец, задачу на экстраполяцию уверенно решают дельфины, хотя ни способ питания, ни весь образ жизни этих животных не создают для этого никаких специальных предпосылок (Флесс и др., 1987).
Особенности врожденного поведения, обеспечивающие кормовую специализацию животных данного вида, не могут быть решающим фактором, обеспечивающим наличие способности к экстраполяции.
Вторым важным подходом к анализу этой проблемы было сопоставление результатов решения тестов, отражающих существование разных форм рассудочной деятельности, помимо способности к экстраполяции. Хищные млекопитаю
Дата добавления: 2021-09-25; просмотров: 687;
Поиск по сайту
Узнать еще
- D. ОСНОВЫ МЕДИЦИНСКОЙ МИКОЛОГИИ
- I. Общая характеристика категории состояния как часть речи
- II. Лексико-грамматические разряды имен числительных. Их характеристика.
- II. Лексико-грамматические разряды местоимений. Их общая характеристика
- II. Методологические основы педагогики.
- II. Основые приемы освоения духовых
- III.Акустические основы настройки музыкальных инструментов
- IV. ОБЩАЯ ХАРАКТЕРИСТИКА ВИРУСОВ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
