Фоторецепторные системы.
Для животных свет – это один из наиболее эффективных видов дистантной рецепции. Живые организмы приспособились к восприятию световых лучей, достигающих поверхности Земли в диапазоне от 200 до 1200 нм.
Фоторецепторные системы появляются уже у кишечнополостных – у медуз. У них фоторецепторные клетки развиваются из эктодермы и сосредотачиваются по краю зонта и на щупальцах. У червей светочувствительные клетки рассеяны по всей поверхности тела, но у некоторых из них на головном конце тела они образуют скопления, окруженные пигментом. Эти, так называемые «глазки», позволяют животным ориентироваться по направлению света. У моллюсков возникает оптический аппарат, превращающий поток света в изображение, что дает начало предметному зрению. Оно совершенствовалось путем эволюции фасеточного и камерного глаза. Кроме того, многие беспозвоночные сохранили рассеянные по поверхности тела светочувствительные клетки.
Реже фоторецепторные клетки лежат на одном уровне с покровным эпителием (например, у медуз). Чаще они погружены в глазную ямку, что предохраняет их от грубых механических раздражений. В ряде случаев поверх рецепторных клеток формируется прозрачная кутикула, обеспечивающая еще более надежную защиту. Если края ямки смыкаются, то образуется пузырчатый (камерный) глаз. В таких случаях прозрачные клетки передней стенки глазного пузырька превращаются в хрусталик, который отбрасывает изображение предметов окружающего мира на фоторецепторные клетки, что дает начало предметному зрению. Большую роль при этом играет пигмент, который изолирует каждую светочувствительную клетку от побочных лучей света за исключением тех, которые падают на нее в строго определенном направлении. Без такой оптической изоляции образное зрение было бы невозможным.
Дальнейшее усовершенствование пузырчатого глаза сводится к увеличению числа фоторецепторных клеток и приобретению диафрагмы – зрачка, устраняющего боковые лучи, а также к появлению аппарата аккомодации, позволяющего глазу настройку на рассмотрение дальних или близких предметов. Аккомодация достигается или перемещением хрусталика или изменением его кривизны.
Глаза камерного типа, где и оптические структуры и фоторецепторные клетки располагаются в одной камере, дают сплошное, но обратное изображение увиденного. Их имеют паукообразные и моллюски, особенно головоногие. Но наибольшего совершенства такие глаза достигают в эволюции позвоночных животных и у человека.
У членистоногих бывают глаза двух типов – простые и сложные (или фасеточные). Простой глаз состоит из единственной линзы, прикрывающей небольшое число фоторецепторных клеток. Такие глаза могут различать свет и темноту, но не способны к предметному зрению.
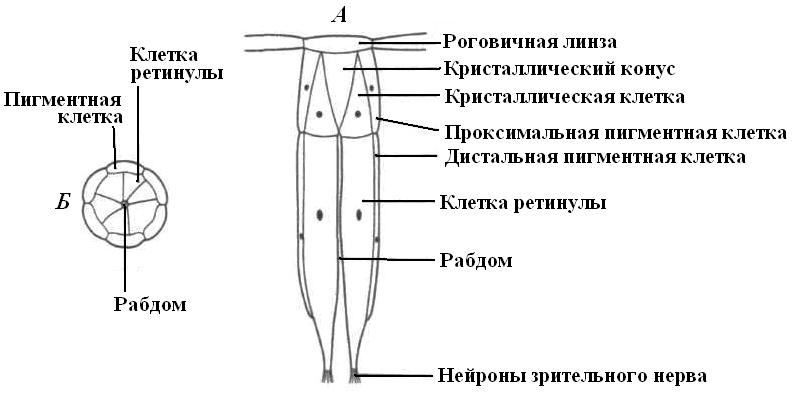
|
| Рис. 23. Схема строения отдельного омматидия. А – продольный разрез; Б – поперечный разрез. |
Фасеточные глаза состоят из тысяч отдельных зрительных единиц – омматидий (рис. 24).
Каждый омматидий имеет две светопреломляющие структуры – роговичную линзу и кристаллический конус, являющийся производным особых клеток. Каждый омматидий обрабатывает изображение только одной точки рассматриваемого предмета. Линза роговицы, образованной прозрачной кутикулой, посылает пучок света на кристаллический конус, где он проходит через отверстие, окруженное пигментными клетками, а затем попадает на фоторецепторные клетки, нервные импульсы от которых попадают в головной ганглий. Изображение прямое, но фасеточное. Фоторецепторы насекомых избирательно чувствительны к тому или иному цвету, но, кроме того, они чувствительны к ультрафиолетовому участку спектра. Например, глаз таракана лучше всего реагирует на зеленый и ультрафиолетовый цвет.
Важным свойством зрения насекомых является способность к прослеживанию быстрых движений, достигающих нескольких сотен в секунду. Фасеточные глаза приспособлены к видению, как на ярком свете, так и в сумерках благодаря особому механизму адаптации фоторецепторов к изменению освещенности.
Камерный глаз имеют некоторые беспозвоночные, однако наибольшего развития глаз такого типа достигает у осьминога, позвоночных и человека.
Глаз состоит из трех оболочек: 1) склеры (фиброзной оболочки), которая в передней части глаза становится прозрачной и образует главную линзу – роговицу; 2) сосудистой оболочки, которая содержит пигмент и представлена собственно сосудистой оболочкой, ресничным телом и радужкой; 3) аккомодационным аппаратом – хрусталиком, обеспечивающим фокусировку изображения на сетчатке; 4) сетчаткой, ретиной содержащей фоторецепторные и нервные клетки. Наполненное водянистой влагой пространство перед радужной оболочкой и хрусталиком называется передней камерой глаза, а позади радужной оболочки – задней. За хрусталиком полость глаза заполнена желеобразным стекловидным телом.
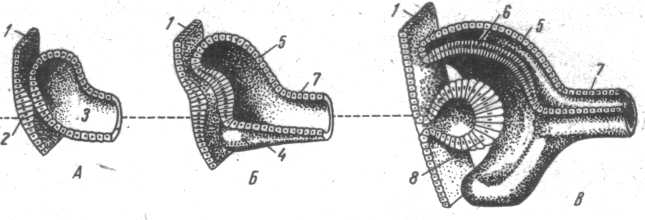
|
| Рис. 24. Развитие глаза. А, Б, В – сагиттальные разрезы эмбрионального зачатка глаза на различных стадиях развития: 1 – эктодерма; 2 – хрусталиковая плакода – будущий хрусталик; 3 – глазной пузырек; 4 – сосудистая выемка; 5 – наружная стенка глазного бокала – будущая сетчатка; 7 – стебелек – будущий зрительный нерв; 8 – хрусталиковый пузырек. |
Эта общая схема строения глаза сохраняется практически во всех группах позвоночных, что отражено в его эмбриональном развитии. Зачаток глаза появляется у эмбриона как пара неглубоких желобков в выростах переднего мозга. Позже эти выросты образуют глазные пузыри, которые связаны с эмбриональным мозгом при помощи глазных стебельков. Глазные пузыри вступают в контакт с эктодермой будущей лицевой части головы и индуцируют в ней развитие хрусталика. Инвагинация стенки глазного пузыря приводит к формированию двухслойного глазного бокала, из наружного слоя которого развивается пигментный слой сетчатки, а из внутреннего – сетчатка. Сосудистая оболочка формируется из окружающей глаз белой мезенхимы. Склера, цилиарная мышца образуются тоже из мезенхимы. Зачаток хрусталика отделяется от эктодермы. Из последней развивается и эпителий роговицы.
Общий план строения глаза не исключает многообразия организации. У многих крокодилов и сумчатых млекопитающих в пигментной оболочке накапливается гуанин, образующий зеркальную прослойку – тапетум. Аналогичная отражающая поверхность имеется у рыб и млекопитающих в сосудистой оболочке. Ее появление связано с необходимостью повышения освещенности фоторецепторного слоя. Свет, который проходит через сетчатку, отражается от зеркальной поверхности тапетума и возвращается к фоторецепторам. Для многих акул и костистых рыб характерно изменение отражающей способности зеркального слоя в зависимости от освещенности.
В глазах рептилий и птиц имеются специфические образования сосудистого происхождения – папиллярный конус и гребень. У рептилий папиллярный конус не связан с фоторецепцией, а участвует в метаболизме внутренней части глаза. Для птиц характерно разнообразное строение гребня, на котором располагается сетчатка.
У позвоночных сетчатка имеет инвертированный характер: ее фоторецепторы расположены на задней дальней от зрачка поверхности сетчатки и развернуты светочувствительными сегментами в сторону пигменного эпителия. Свет проходит через всю толщу сетчатки и тела самих фоторецепторов, прежде чем попадет на зрительный пигмент мембраны дисков дистальных отростков фоторецепторных клеток.
У многих млекопитающих глаза расположены фронтально, чем достигается бинокулярное зрение. В таких случаях зрительные поля обоих глаз перекрываются, что имеет ряд преимуществ, по сравнению с использованием каждого глаза в отдельности. Чем больше глаза направлены вперед, тем шире поле стереоскопического зрения. У человека, например, общее поле зрения охватывает 180°, а стереоскопическое – 140°. У животных, у которых глаза расположены по бокам головы, фронтальное стереоскопическое поле бывает ограниченным и используется лишь для рассмотрения удаленных предметов. Для того, чтобы лучше рассмотреть близкий предмет, животное поворачивает голову и использует монокулярное зрение.
Стереоскопическое зрение позволяет получить более точное представление о размерах и форме предметов, а также о расстоянии, на котором они находятся. В основном стереоскопическое зрение характерно для хищных животных. У животных, которым приходится спасаться от хищников, глаза, наоборот располагаются по бокам головы, благодаря чему они имеют более широкий обзор, но ограниченное стереоскопическое зрение, так, например, у кролика общее поле зрение составляет 360°, а стереоскопическое – всего 20°.
Дата добавления: 2019-12-09; просмотров: 1041;
Поиск по сайту
Узнать еще
- I. История возникновения и развития классно-урочной системы.
- III. Филогенез эндокринной системы.
- А) Реформа политической системы.
- Автоматизированные загрузочные устройства – важнейшая часть механообрабатывающей системы. Классификация устройств загрузки . Питатели.
- Анатомо-физиологические особенности сердечно-сосудистой системы. (АФО с.с.с) Анатомия сердца.
- Ассоциативные системы.
- Балочные системы. Определение реакций опор и моментов защемления
- Биотическая структура экосистемы.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
