Органы боковой линии.
У рыб и живущих в воде амфибий присутствует особый вид механорецепторов – органы боковой линии. Они представляют собой отдельные ямки или систему каналов, которые идут вдоль боковой поверхности тела в виде полоски.
И каналы и ямки сообщаются с внешней средой порами. В каналах и на дне ямок расположены чувствительные бугорки – нейромасты. В некоторых случаях (например у шпорцевой лягушки) последние лежат прямо на поверхности тела. В нейромастах имеются волосковые клетки, относящиеся к безотростковым механорецепторам. Волоски этих клеток погружены в студенистые бугорки, которые называются купулами. На поверхности рецепторных клеток имеются два ряда ресничек, одна из которых неподвижная – киноцилия, а другая тонкая подвижная – стереоцилия. Если стереоцилии отклоняются в сторону киноцилий, волосковая клетка деполяризуется, если же стереоцилии отклоняются в противоположную от киноцилий сторону, то волосковая клетка гиперполяризуется. В нейромастах волосковые клетки располагаются попарно, чем достигается различение направленности движения воды. К каждой волосковой клетке подходят эфферентное и афферентное нервные волокна, чем обеспечивается дискретность информации.
Органы равновесия.
Механорецепция как средство ориентации в пространстве наблюдается уже у простейших. Инфузории различают «верх» и «низ», используя твердые внутриклеточные включения как прототипы статоцистов высших животных. У медуз по краю колокола располагаются статоцисты – замкнутые пузырьки, в полости которых статолиты, состоящие из CaSO4 , своей тяжестью раздражают первичные рецепторные клетки, сигнализируя о положении тела в пространстве. У ракообразных в статоцистах, расположенных у основания антеннул, образуются вертикальный и горизонтальный каналы, движение жидкости в которых раздражает волоски групп клеток, осуществляющих рецепцию углового ускорения. Роль статолитов в них играют песчинки.
У позвоночных основную роль в определении положения и движения тела играет вестибулярный аппарат. Он развивается на основе органа боковой линии рыб, из полости которого выпячиваются мешочки, дающие начало полукружным каналам. У миксин образуется единственный вертикальный канал, у миног их два, а у остальных их три. Они дают информацию о движениях в любом направлении трехмерного пространства.
Вестибулярный аппарат высших позвоночных животных представлен двумя мешочками и тремя полукружными каналами с ампулами. Один из мешочков круглый (саккулюс), другой – овальный (утрикулюс). В них имеются участки с сенсорными (чувствительными) волосковыми клетками, которые входят в состав, так называемых макул. В последних присутствуют еще и опорные клетки. Поверх таких скоплений клеток располагается студенистая отолитовая мембрана с вкрапленными в нее кристаллами карбоната кальция – отолитами. На верхней поверхности сенсорных клеток имеется кутикула, от которой отходят неподвижные чувствительные волоски – стереоцилии и одна подвижная ресничка – киноцилия, располагающаяся полярно относительно пучка стереоцилий.
Стимуляция сенсорных клеток обеспечивается скольжением по их поверхности отолитовой мембраны. Возникающее при этом возбуждение воспринимается чувствительными нейронами, дендриты которых образуют синапсы на сенсорных клетках. Возникающий при этом нервный импульс передается в соответствующую часть вестибулярного анализатора.
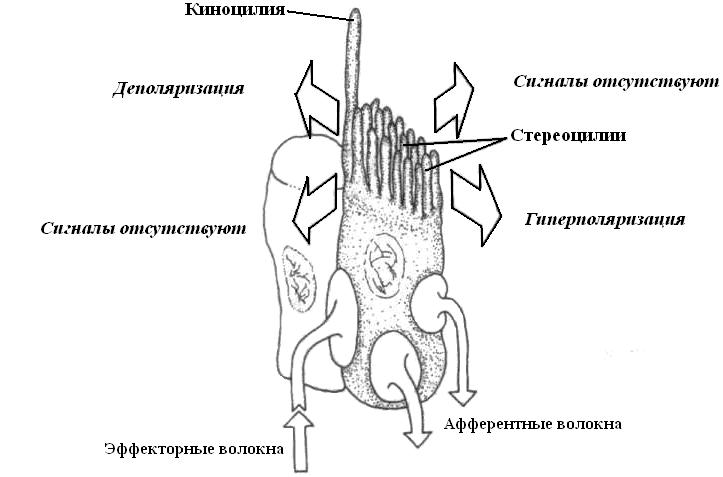
|
| Рис. 18. Строение и принципы чувствительности волосковых механорецепторов в органах боковой линии, слуховой и вестибулярной системах. |
Макулы овальных мешочков – это места восприятия линейных ускорений, рецепторы гравитации (земного притяжения), связанные с изменением тонуса мышц, определяющих установку тела. Макулы круглых мешочков, кроме гравитации, воспринимают и вибрационные колебания.
Рецепторные образования вестибулярного аппарата присутствуют и в ампулах полукружных каналов (рис. 19).

|
| Рис. 19. Схема строения ампулы полукружного канала (поперечный разрез). |
Они воспринимают сигналы главным образом о движении головы в трех взаимоперпендикулярных направлениях и называются кристами (ампулярными гребнями). Кристы имеют вид поперечных складок, состоящих из волосковых и поддерживающих эпителиальных клеток, над вершинами которых располагается желеобразный купол. Последний смещается при движении эндолимфы в полукружных каналах и раздражает волосковые клетки. Их возбуждение воспринимается волокнами вестибулярного нерва и передается в соответствующие участки головного мозга, откуда нервные импульсы поступают скелетным мышцам.
Орган слуха.
Приспособление механорецепторов к восприятию звуков в ходе эволюции было связано не только с появлением специфических звуковоспринимающих клеток, но и вспомогательных структур, позволяющих дифференцировано реагировать на звуки. В этом отношении животные обнаруживают большое разнообразие, связанное с их экологией.
Среди беспозвоночных органы слуха имеются у членистоногих. Они служат для них органами коммуникации, а в ряде случаев позволяют спасаться от врагов. У одних органы слуха представлены тимпанальными мембранами, у других – хордотональными слуховыми органами.
Тимпанальные мембраны натянуты на прочной хитиновой рамке и выполняют роль барабанной перепонки. Под мембраной лежит трахейный пузырь, а между ним и мембраной – группы рецепторных клеток.
Хордотимпальные слуховые органы состоят из струнообразной связки, натянутой над покровами тела, и сенсорных клеток, которые одним концом соединяются со связкой, а другим – с покровами. Связка колеблется в унисон звуковым волнам, поступающим из окружающей среды, и возбуждает рецепторные клетки.
Орган слуха позвоночных развивается вместе с органом равновесия из утолщения эктодермы в задней части головы. Этот зачаток углубляется и принимает вид пузырька, который в дальнейшем разделяется на 2 отдела. Из верхнего отдела образуются полукружные каналы и овальный мешочек, а из нижнего формируется полое выпячивание – лагена и круглый мешочек. Все эти образования заполнены жидкостью – эндолимфой и составляют так называемый перепончатый лабиринт. Он окружен стенками скелета, а промежуток между ними и перепончатым лабиринтом заполнен перилимфой.
У круглоротых, рыб и безногих амфибий со слуховой рецепцией связаны 2 сосочка круглого мешочка. Для амфибий характерен амфибийный сосочек, а для остальных тетрапод – базилярный сосочек, в которых звуковоспринимающую функцию выполняют рецепторные волосковые клетки.
У рептилий, птиц и однопроходных имеется вытянутый слепой вырост – лагена. У птиц она к тому же изгибается, а у млекопитающих многократно закручивается, образуя улитку. У млекопитающих литка разделяется тремя перепонками на 3 части.
Две из них – это лестница преддверия и барабанная лестница, содержат перилимфу и соединяются между собой на верхушке улитки через небольшое отверстие. Средний канал улитки (перепончатый лабиринт) содержит эндолимфу. Он отделен от барабанной лестницы базилярной мембраной, а от лестницы преддверия – рейснеровой. Базиллярная мембрана состоит из поперечно ориентированных коллагеновых волокон («струн»), длина которых по направлению к основанию улитки постепенно уменьшается. Эти волокна резонируют в унисон звукам различной высоты.
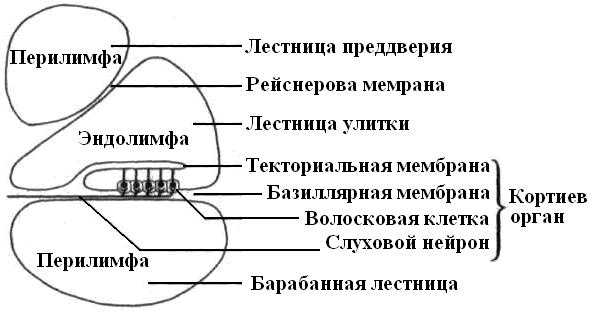
|
| Рис. 20. Схема поперечного разреза улитки, на котором виден кортиев орган. |
Располагающийся на базилярной мембране звуковоспринимающий кортиев орган состоит из сенсорных волосковых и поддерживающих эпителиальных клеток, которые подразделяются на наружные и внутренние.
На апикальных участках сенсорных клеток имеются пучки коротких уплотненных микроворсинок, способных отклоняться – это стереоцилии. Стереоцилии наружных сенсорных клеток своими вершинами прикасаются к поверхности лежащей над ними текториальной мембраны. Наружные сенсорные эпителиоциты значительно чувствительнее к звукам большей интенсивности, чем внутренние.
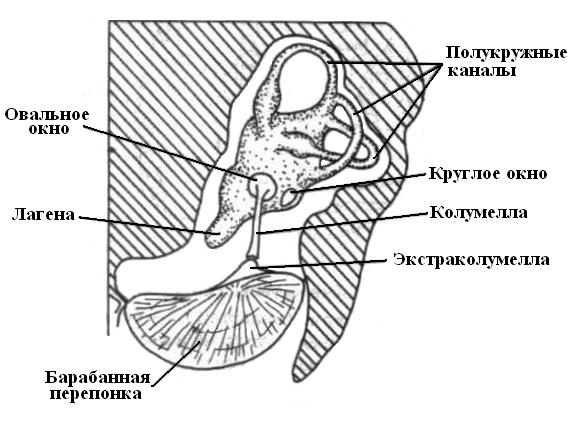
|
| Рис. 21. Организация слуховой системы птиц. |
Первичноводные позвоночные довольно широко используют коммуникации, связанные с испусканием звуков. Очень часто они для этого используют растяжение плавательного пузыря. Звуковоспринимающую функцию у них выполняют рецепторные клетки макул круглого мешочка и лагены. Звуковые волны эффективно передаются в воде, поэтому у первичноводных животных нет специализированных образований для фильтра и усиления звуков. Эту функцию выполняет все тело, по тканям которого колебания воды передаются практически без потерь.
У наземных позвоночных условия звукопередачи на внутреннее ухо изменяются, т.к. воздействие звуковой волны оказывается слишком слабым для рецепторного аппарата внутреннего уха. Поэтому у них в эволюции сформировались специальные приспособления для усиления звуковых сигналов – среднее и наружное ухо. Все особенности структур среднего уха направлены на усиление внешних звуковых сигналов.
Вся система среднего уха сложилась на основе морфогенетических изменений одной из висцеральных щелей – брызгальца, наружная часть которого несколько расширилась, образовав барабанную полость, и закрылась барабанной перепонкой, а внутренняя сузилась и превратилась в евстахиеву трубу – узкий канал, конец которого, как и брызгальца открывается в глотку. В среднем ухе имеется набор косточек, которые передают колебания от барабанной перепонки к овальному окну. У амфибий, рептилий и птиц сигналы передаются через столбик (колумелла), а у млекопитающих – через наковальню, молоточек и стремечко.
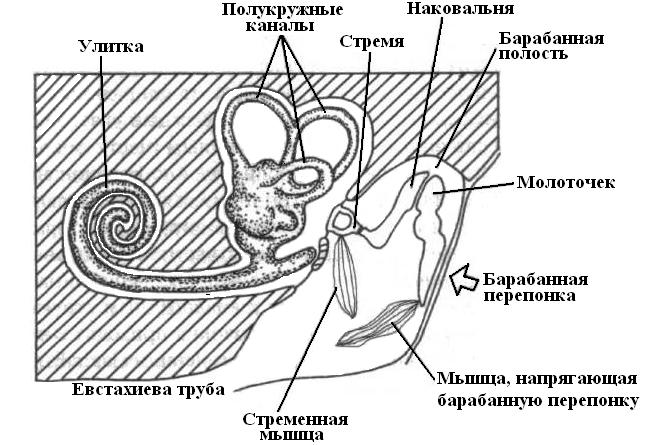
|
| Рис. 22. Организация слуховой системы млекопитающих. |
У змей и некоторых других рептилий барабанная перепонка редуцирована. У них колебания воздуха передаются от квадратной черепа через короткую связку колумелле, а от нее к овальному окну.
Наружное ухо присуще птицам и млекопитающим. Птицы обладают слухом в большом частотном диапазоне, что обеспечивается организацией рецепторного аппарата и особенностями наружного уха. У птиц лагена в сравнениями с рептилиями увеличена в размерах, но еще не формирует завитков, как у млекопитающих. Слуховые рецепторные клетки у птиц располагаются в лагене в перепончатом лабиринте, который во многом похож на лабиринт млекопитающих.
У млекопитающих слуховой аппарат внутреннего уха представляет собой длинный рукав, который закручен в виде улитки. Исключение составляют однопроходные (яйцекладущие), у которых практически прямой рукав внутреннего уха (нет улитки). Полость среднего уха соединяется с ротовой полостью евстахиевой трубой и отличается главным образом большой сложностью передаточного аппарата, состоящего из трех слуховых косточек. Особая мышца, прикрепляющаяся к рукоятке молоточка, натягивает барабанную перепонку, а другая – мышца стремени, наклоняет стремечко и натягивает перепонку овального окна. Рефлекторно сокращением этих мышщ уменьшается амплитуда колебаний всего аппарата при более сильных звуках.
У млекопитающих значительного развития достигает наружный слуховой проход и связанные с ним наружные ушные раковины. Последние собирают звуковые волны и направляют их в проход. Ушная раковина имеет хрящевой скелет и приводится в движение особой мускулатурой. У китообразных в связи с их образом жизни ушная раковина отсутствует, наружный слуховой проход редуцирован, и звуки проводятся к среднему и внутреннему уху через жировые и костные ткани черепа.
Одной из основных характеристик слуха является восприятия звуков определенного диапазона частот. Наибольшее разнообразие в этом отношении характерно для позвоночных животных. Так, рыбы чутко реагируют на инфразвуковые и низкочастотные колебания, издаваемые плавающими в воде животными. Сами рыбы издают с аналогичным диапазоном звуки типа: хрюканья, крика, тарахтенья, гудков, скрежета и др. Пресмыкающиеся слышат звуки более высокого тона, чем амфибии. Большинство птиц наиболее чувствительны к звукам 3,0 – 3,5 кГц, собаки – 35 кГц, летучие мыши и дельфины до 100 кГц, человек от 16 до 18 кГц. В то же время тимпанальные органы кузнечиков слышат звуки около 200 кГц.
Согласованная деятельность обоих ушей обеспечивает по разности силы и фазовому сдвигу звуковых колебаний, приходящих в одно и другое ухо, определять направление к источнику звука.
Дата добавления: 2019-12-09; просмотров: 1609;
Поиск по сайту
Узнать еще
- F45.38 другие органы или системы
- V. Органы осуществляющие правосудие
- Амиотрофический боковой склероз
- Боковой удар левой в голову с шагом назад левой ногой.
- Боковой удар левой в туловище с шагом влево.
- Введение поправок в измеренную длину линии. Определение неприступных расстояний. Дальномеры
- Вегетативные органы растений.
- Вегетативные органы растений. Их трансформация в

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
