Сайт 0-репликации (ориджин репликации, сайт инициации репликации, точка инициации репликации)
Этот процесс продолжается непрерывно по длине копируемой структуры и заканчивается в этом же репликоне образованием двух молекул «полуконсервативного» типа.
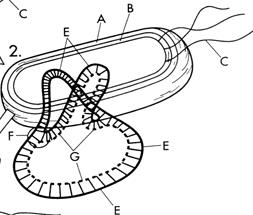
А- клеточная стенка бактерии; В –мембрана бактериальной клетки; С- жгутики; D-цитоплазма; F- энзим; Е – исходная ДНК; G- новая ДНК; О - место начала репликации.
В больших линейных молекулах ДНК эукариот имеется много точек начала репликации и соответствующих им репликонов (от нескольких сотен до десятков тысяч), т. е. такая ДНК является полирепликонной.
При рассмотрении современных представлений о механизме репликации ДНК эукариот можно условно выделить три последовательных этапа этого процесса, происходящего в репликоне, в каждом из которых принимают участие те или иные белки (ферменты).
Первый этап связан с быстрым раскручиванием двух полинуклеотидных нитей спирализованной молекулы ДНК на определенном ее участке (в границах работающего репликона) и с их разделением путем разрушения водородных связей между парами комплементарных оснований. При этом образуются два одноцепочечных фрагмента родительской молекулы, каждый из которых может выступать в роли матрицы для синтеза комплементарной (дочерней) нити. Этот этап инициируется в соответствующей точке начала репликации и обеспечивается комплексным участием нескольких различных белков. В результате их действия формируется T-образная структура, названная вилкой репликации, в которой две родительские цепочки ДНК уже отделены друг от друга (рис.). Образовавшаяся вилка репликации быстро продвигается вдоль двойной спирали родительской молекулы ДНК благодаря активности «расплетающего» фермента ДНК-хеликазы и при участии группы дестабилизирующих белков.
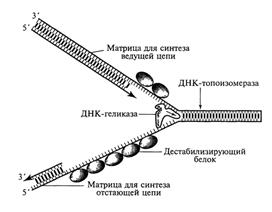
Рис. Схема образования репликационной вилки ДНК
Эти белки обладают способностью связываться только с одноцепочечными (уже раскрученными и разделенными) участками молекулы, препятствуя возникновению на них вторичных складчатых образований («шпилек») за счет случайных соединений между комплементарными нуклеотидами однонитевой структуры. Следовательно, они способствуют выпрямлению однонитевых участков молекулы, что необходимо для нормального выполнения ими матричных функций.
Быстрое расплетание ДНК с помощью хеликазы без дополнительного вращения нитей по отношению друг к другу должно приводить к образованию новых витков (узлов) на участках родительской молекулы перед движущейся вилкой репликации, создающих повышенное топологическое напряжение на этих участках. Такое напряжение устраняется еще одним белком (ДНК-топоизомеразой), который, перемещаясь вдоль двухспиральной родительской ДНК перед вилкой репликации, вызывает временные разрывы в одной из цепочек молекулы, разрушая фосфодиэфирные связи и присоединяясь к разорванному концу. Возникший разрыв обеспечивает последующее вращение нити двойной спирали, что, в свою очередь, приводит к расплетанию образующихся супервитков (узлов). Поскольку разрыв полинуклеотидной цепочки, вызванный топоизомеразой, носит обратимый характер, то разорванные концы быстро воссоединяются сразу после разрушения комплекса этого белка с разорванным концом.
На втором этапе происходит матричный синтез новых (дочерних) полинуклеотидных цепей на основе известного принципа комплементарного соответствия нуклеотидов старой (матричной) и новой цепей. Этот процесс осуществляется путем соединения (полимеризации) нуклеотидов новой цепи с помощью ферментов ДНК-полимераз нескольких типов. Следует отметить, что ни одна из известных сегодня ДНК-полимераз не способна начать синтез нового полинуклеотида путем простого соединения двух свободных нуклеотидов. Инициация этого процесса требует наличия свободного 3'-конца какой-либо полинуклеотидной цепочки ДНК (либо РНК), которая соединена с другой (комплементарной) цепочкой ДНК. Иными словами, ДНК-полимераза способна лишь добавлять новые нуклеотиды к свободному 3'-концу имеющегося полинуклеотида и, следовательно, способна наращивать эту структуру только в направлении 5'→3'.
С учетом указанного обстоятельства становится понятным асимметричный характер функционирования вилки репликации (рис.). Как видно из приведенных схем, на одной из матричных нитей вилки 3'→5') идет относительно быстрый и непрерывный синтез дочерней нити (ведущей, или лидирующей, цепочки) в направлении 5' →3', тогда как на другой матрице (5'→ 3') идет более медленный и прерывистый синтез отстающей цепочки короткими фрагментами (100-200 нуклеотидов), получившими название фрагментов Оказаки, и также в направлении 5'→ 3'.
Синтез ведущей и отстающей цепочек осуществляют ДНК-полимеразы разных типов. Свободный 3'-конец, необходимый для начала синтеза фрагмента Оказаки, обеспечивается короткой нитью РНК (около 10 нуклеотидов), получившей название РНК-праймера (РНК-затравки), которая синтезируется с помощью фермента РНК-праймазы. РНК-праймеры могут комплементарно спариваться сразу с несколькими участками на матричной нити ДНК, создавая условия для одновременного синтеза нескольких фрагментов Оказаки при участии ДНК-полимеразы III (рис. 1.10). Когда синтезированный фрагмент Оказаки достигает 5'-конца очередного РНК-праймера, начинает проявляться 5'-экзонуклеазная активность ДНК-полимеразы I, которая последовательно выщепляет нуклеотиды РНК в направлении 5'→ 3'. При этом удаляемый РНК-праймер замещается соответствующим фрагментом ДНК.
Последний (третий) этап рассматриваемого процесса связан с действием фермента ДНК-лигазы, который соединяет 3'-конец одного из фрагментов Оказаки с 5'-концом соседнего фрагмента с образованием фосфодиэфирной связи, восстанавливая таким образом первичную структуру отстающей цепочки, синтезируемой в функционирующем репликоне. Дальнейшая спирализация появившегося «полуконсервативного» участка ДНК (закручивание спирали) происходит с участием ДНК-гиразы и некоторых других белков.
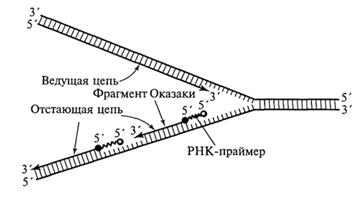
Рис. Синтез ведущей и отстающей цепей ДНК в области репликационной вилки
Полирепликонный принцип организации молекулы ДНК различных эукариот, в том числе человека, обеспечивает возможность последовательного копирования генетического материала этих организмов без одновременного раскручивания (деспирализации) всей огромной по размерам и сложно упакованной молекулы, что значительно сокращает время ее репликации. Иными словами, в тот или иной момент времени в одной группе репликонов молекулы процесс копирования может быть уже завершен объединением и спирализацией соответствующих участков, тогда как в другой группе он только начинается расплетанием двухнитевых структур.

Рис. Схематическое изображение процесса репликации, цифрами обозначены: 1- запаздывающая цепь, 2 - цепь-лидер, 3- ДНК-полимераза (Polα), 4- ДНК лигаза, 5- РНК-праймер, 6- ДНК- праймаза, 7- фрагмент Оказаки, 8- ДНК-полимераза (Polδ), 9- хеликаза, 10- одиночная цепь со связанными белками, 11- топоизомераза
Ферменты, участвующие в репликации ДНК эукариот:
1.ДНК-хеликаза и дестабилизирующие белки; они расплетают двойную спираль родительской ДНК и формируют репликационную вилку.
2.ДНК-полимеразы, которые катализируют синтез полинуклеотидной цепи ДНК в направлении 3'-5, копируя в репликационной вилке матрицу с высокой степенью точности. Поскольку две цепи двойной спирали ДНК антипараллельны, в направлении 5'-3' непрерывно синтезируется лишь одна из двух цепей - ведущая; другая цепь - отстающая, синтезируется в виде коротких фрагментов Оказаки. ДНК-полимераза способна к исправлению собственных ошибок, но не может самостоятельно начать синтез новой цепи.
3.ДНК-праймаза, которая катализирует короткие молекулы РНК-затравки. Впоследствии фрагменты РНК удаляются - их заменяет ДНК.
4.Теломераза, заканчивающая построение недорепликацированых 3'-концов линейных молекул ДНК.
5. ДНК-топоизомеразы, помогающие решить проблемы кручения и спутывания спирали ДНК.
6. Инициаторные белки, связывающиеся в точке начала репликации и
способствующие образованию нового репликационного глазка с одной или двумя вилками. В каждой из вилок вслед за инициаторными белками к расплетенной ДНК сначала присоединяется белковый комплекс, состоящий из ДНК-хеликазы и ДНК-праймазы (праймосома). Затем к праймосоме добавляются другие белки и возникает «репликационная машина», которая и осуществляет синтез ДНК.
Дата добавления: 2017-04-05; просмотров: 2249;
Поиск по сайту
Узнать еще
- Web-сайт организации
- Ізопотенційна точка електродів
- Билет № 35. Точка зрения и ее разновидности. Смена «точки зрения» и композиция эпического произведения. М.М.Бахтин о романном разноречии.
- В качестве примера рассмотрим создание бесплатного сайта на OKIS
- Веточка раскаленного железа
- Визначення динамічних реакцій в точках закріплення осі тіла, що обертається
- ВИЗНАЧЕННЯ ПОЛОЖЕНЬ ЛАНОК ГРУП І ПОБУДОВА ТРАЄКТОРІЙ, ОПИСУВАНИХ ТОЧКАМИ ЛАНОК МЕХАНІЗМУ.
- Визначення реактивних ударних імпульсів в точках закріплення осі

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
