Фотосинтез у зеленых растений
Энергия фотонов преобразуется в энергию химических связей синтезируемых органических веществ.
Скорость фотосинтеза измеряется количеством кислорода, выделенного за единицу времени. Она зависит от интенсивности поглощенного света. Максимальная скорость фотосинтеза при постоянном сильном освещении достигает одной молекулы 02 (на одну молекулу хлорофилла) за 50 с. Источником всего атмосферного кислорода служит вода, участвующая в реакциях фотосинтеза.
При восстановлении углекислого газа до углевода осуществляется перенос 4 атомов водорода с Н20 на С02. Изменение свободной энергии в реакциях фотосинтеза при образовании одной молекулы 02 составляет около 500 кДжмоль1, на что затрачивается 8 квантов света с суммарной энергией примерно 1470 кДж моль1. Следовательно, коэффициент использования солнечной энергии при фотосинтезе достигает 34%. Примерно к такому же КПД можно прийти путем другого расчета — при сопоставлении энергии, заключенной в 8 фотонах, с энергией, затрачиваемой на синтез углеводов.
Свободная энергия, затрачиваемая на транспорт электронов против разности потенциалов при переносе атомов водорода с Н20 на С02 черпается из фотохимических процессов, в которых участвует хлорофилл. Каждая молекула хлорофилла тесно связана с донором (Д) и акцептором (А) электронов. Поэтому при ее фотовозбуждении происходят следующие превращения. Акцептор принимает электрон от возбужденного хлорофилла и превращается в А-. Место этого электрона в хлорофилле занимает другой, принимаемый им от донора. Электрон не может перейти с донора на акцептор непосредственно, так как они пространственно разобщены и не взаимодействуют между собой. Пространственное разобщение их обеспечивается тем, что донор и акцептор являются компонентами биологической мембраны и фиксированы в ее определенных точках. Не взаимодействуя между собой, А' и Д+, образовавшиеся под действием света при участии хлорофилла, могут участвовать в других окислительно-восстановительных реакциях с теми мембранными компонентами, которые тесно контактируют с ними. Вне биомембраны фотосинтез не осуществляется.
Местом фотосинтеза в зеленых растениях являются мембранные системы хлоропластов. В этом специализированном органоиде, кроме двойной наружной мембраны, содержится сложно организованная сеть внутренних мембран, образующих многослойные структуры, упакованные в пачки (граны). Внутренние мембраны ограничивают замкнутые объемы в виде уплощенных пузырьков — тилакоидов. В каждом хлоропласте примерно 1000 ти- лакоидов, а в одной клетке - от 50 до 200 хлоропластов. В мембранах тилакоидов протекают светозависимые фотосинтетические реакции, так как там сосредоточены хромофоры — молекулы, поглощающие световую энергию. К ним относятся хлорофилл а, хлорофилл Ь, каротиноиды и другие пигменты. Хлорофилл поглощает свет наиболее интенсивно в синей и красной, но отражает в зеленой и желтой областях. Этим определяется зеленый цвет растений. На один хлоропласт приходится примерно 109 молекул хлорофилла.
Две разновидности хлорофилла (а и Ь), имея незначительные различия в химическом строении, существенно отличаются по физическим свойствам и биологической активности. О коренных отличиях в механизмах переноса энергии свидетельствует наличие флуоресценции у хлорофилла а (с квантовым выходом около 0,2) при ее отсутствии у хлорофилла. Последний сам по себе непосредственно не участвует в фотосинтетических превращениях, но он входит в состав светособирающей пигментной матрицы, помогающей улавливать солнечную энергию. После фотовозбуждения хлорофилл b быстро передает свою избыточную энергию хлорофиллу а, который при этом возбуждается и принимает непосредственное участие в фотосинтезе. Хлорофилл а может возбуждаться светом и без посредника, но участие хлорофилла b повышает коэффициент использования солнечной энергии. Он повышается также благодаря присутствию в светособирающей матрице других пигментов (каротиноидов и фикобилинов). Сопоставив спектры поглощения хлорофилла и каротиноидов, нетрудно убедиться, что хлорофилл поглощает энергию ограниченного участка солнечного спектра. Однако фотосинтез идет под действием почти всей видимой области электромагнитного спектра, поскольку зеленый свет хорошо поглощают каротиноиды, входящие в светособирающую матрицу. Кроме того, каротиноиды защищают хлорофилл от фотоокисления при избыточном освещении.
Перенос энергии от различных пигментов этой матрицы на хлорофилл а осуществляется по резонансному механизму. В конце концов, энергия фотовозбуждения достигает реакционного центра, в котором сосредоточено несколько молекул хлорофилла а, ориентированных определенным образом в тилакоидной мембране. Здесь начинается первая стадия фотохимического процесса. Совокупность реакционного центра и светособирающей матрицы называется фотосинтетической единицей. Каждая из них включает приблизительно 300 молекул хлорофилла.
Фотосинтетическая единица входит в состав фотосистемы (ФС), компонентами которой являются также интегральные белки тилакоидной мембраны. Мембранные протеины фиксируют многие молекулы хлорофилла на одной полипептидной цепи, создавая тем самым необходимую ориентировку пигмента. Комплекс «хлорофилл а — мембранный белок» служит главным узлом фотогенератора, как иногда называют фотосистему. Функциональным элементом тилакоидной мембраны считается квантосома, в состав которой входят две фотосистемы, не одинаковые по свойствам. В каждой ФС есть собственная фотосинтетическая единица, причем относительное содержание хлорофилла а в ФС I много выше, чем в ФС II. Обе фотосистемы поглощают свет с длинами волн короче 680 нм, но только ФС I способна возбуждаться и более длинноволновым светом, причем ее максимум поглощения приходится на 700 нм. Поэтому хлорофилл а, входящий в разные фотосистемы, имеет различное наименование: в ФС I его называют пигментом Р700, а в ФС II—пигментом Р680. Различия определяются не особенностями молекулярной структуры пигментов (она одинакова), а специфическим мембранным окружением хлорофилла а в ФС I и ФС II (вероятно, составом мембранных белков, фиксирующих молекулы пигмента). В целом молекулярная масса квантосомы достигает 2106 дальтон. Между молекулами хлорофилла, принадлежащими разным ФС в одной квантосоме, расположены молекулы других веществ, входящих в ее состав и выполняющих функцию переноса электронов. Поэтому говорят, что ФС I и ФС II разобщены в пространстве тилакоидной мембраны, но сопряжены между собой системой переноса электронов, которые переносятся с ФС II на ФС I, и обратный переход невозможен.
В темноте молекула хлорофилла а пребывает на нижнем синглетном уровне. Световая энергия по резонансному механизму переводит его электроны на более высокий энергетический уровень. Полосы поглощения хлорофилла соответствуют переходам S —►Sj* (в красной области спектра) и Sq—>S2* (в синей области). Раствор хлорофилла a in vitro флуоресцирует за счет обратного перехода электронов с S* на S0 (заметим, что обратный переход с S2* на S , — безызлучательный). С уровня S2* возможен также безызлучательный переход на метастабильный триплетный уровень Т. Находясь на нем, электроны хлорофилла могут перейти под действием света на уровень Т*, что означает возбужденное триплетное состояние хлорофилла. Именно такой переход присущ хлорофиллу in vivo, когда он встроен в тилакоидную мембрану. С уровня Т электроны могут возвратиться и на уровень S0, что сопровождается фотолюминесценцией. Поэтому хлорофилл а люминесцирует не только в пробирке, но и в хлоропластах. Однако его квантовый выход in vivo только 0,03-0,06, тогда как in vitro — 0,2. Из сопоставления этих чисел ясно, что львиная доля энергии, выделяющейся при электронных переходах в облученной молекуле хлорофилла а, находящейся в тилакоидной мембране, идет на фотосинтез.
Фотосинтез осуществляется за счет перехода возбужденных электронов с Т на Т*, при котором изменяется их спин. Появление же неспаренных электронов приводит к образованию свободных радикалов. Возникновение свободных радикалов при фотосинтезе доказано методом ЭПР.
Полный цикл фотосинтетических превращений начинается с фотовозбуждения хлорофилла а в ФС II (Р680). Возбужденный электрон переносится с Р680 на акцептор, тесно контактирующий с хлорофиллом в тилакоидной мембране. По-видимому, акцептором служит цитохром 555 (Q). На переход затрачивается энергия возбуждения электрона, так как он преодолевает восстановительный потенциал в 1,24 В (идет «в гору»). Приняв электрон, акцептор восстанавливается, а в молекуле хлорофилла, отдавшей его, возникает дырка. Она заполняется электроном воды, которая расщепляется в процессе фотосинтеза с образованием кислорода, протонов и электронов. По-видимому, расщепление воды (точнее, ее окисление до 02) производит не сам возбужденный хлорофилл, а некое промежуточное окисленное соединение, образующееся при возбуждении Р680. Окисление воды, приводящее к выделению кислорода, катализируется ферментной системой, содержащей трехвалентный ион марганца.
Присоединив электрон, хлорофилл (Р680) восстанавливается, и к нему возвращается способность возбуждаться новыми квантами света. Кислород выделяется в атмосферу. Протоны закисляют содержимое тилакоидов, поддерживая там более высокую концентрацию Н+ по сравнению с цитозолем. Такова судьба всех продуктов расщепления воды при фотосинтезе.
Теперь следует обратиться к электронам, покинувшим молекулы хлорофилла (Р680) и восстановившим вещество Q. Проследим их миграцию по цепи переноса ФС II. С вещества Q они последовательно переходят от одного компонента транспортной цепи к другому в направлении более высокого положительного восстановительного потенциала («с горы») — к пластохинону и далее к пластоцианину. Свободная энергия, выделяющаяся при этом ступенчато (дискретно), запасается тилакоидной мембраной и обеспечивает синтез АТФ из АДФ и ортофосфата. Так как скорость переноса электронов по транспортной цепи ФС II не зависит от температуры, то возникло предположение о туннельном механизме миграции электронов между окислительно-восстановительными компонентами этого каскада. Последним из них в ФС II является Р700, т. е. хлорофилл а, принадлежащий другой фотосистеме (ФС I) в данной квантосоме.
Однако Р?00 приобретает способность принять электроны из ФС II только после того, как сам поглотит свет, возбудится и потеряет свой электрон, который поступит в транспортную цепь ФС I. При потере электрона в молекуле Р700 возникает дырка, которую и занимает электрон, пришедший сюда из ФС II (с пластохинона). Следовательно, возбужденный хлорофилл, входящий в состав ФС II, восстанавливает пластохинон (до гидрохинона), а возбужденный хлорофилл, принадлежащий ФС I, окисляет гидрохинон до пластохинона, возвращая ему способность принимать новые электроны от Р680. Такое сопряжение в работе двух фотосистем осуществляется в пределах одной квантосомы. В ней между молекулами хлорофилла, принадлежащими разным фотосистемам, иными словами, между Р680 и Р700, расположено примерно 5 молекул пластохинона. Пигмент Р?00, приняв электроны от компонентов ФС II, восстанавливается, и к нему возвращается способность возбуждаться светом.
В ФС I пока не идентифицировано вещество с наибольшим (по модулю) отрицательным восстановительным потенциалом, на которое переходят возбужденные электроны с Р700. Обозначим его буквой X. Приняв электрон от возбужденного хлорофилла, X становится сильным восстановителем (X), с которого начинается второй каскад переноса электрона в сторону более высокого положительного восстановительного потенциала. Последним компонентом цепи транспорта электронов в ФС I является НАДФ+, восстанавливаемый до НАДФН. Донором водорода служит вода, расщепившаяся под действием света при участии хлорофилла (Р680) еще на первой стадии фотосинтеза.
Конечным акцептором электронов, прошедших последовательно ФС II и ФС I, является С02. Перенос на него электронов с редокс-пары НАДФ+/ НАДФН (восстановительный потенциал равен -0,324 В) осуществляется сложной ферментной системой, входящей в так называемый цикл Кальвина — последовательность химических реакций, протекающих в 13 стадий. Энергия, выделяемая при транспорте электронов с названной редокс-пары на С02, недостаточна для его восстановления до углеводов. Дополнительная энергия, необходимая для синтеза углеводов, поступает в цикл Кальвина в форме АТФ. Так, образование одной молекулы гексозы из 6 молекул С02 обеспечивается не только переносом электронов с НАДФН на С02, но еще и 18 молекулами АТФ. Этот макроэрг образуется также в реакциях фотосинтеза посредством преобразования солнечной энергии, поглощенной хлорофиллом. Синтезу АТФ служит транспорт в квантосоме (в ФС II и ФС I) до этапа восстановления НАДФ+.
За счет поглощения двух квантов света двумя молекулами хлорофилла (Р680 и Р700), принадлежащими одной квантосоме, происходит перенос одного электрона на С02. Кроме того, энергия этих двух фотонов обеспечивает перенос одного электрона с воды на Р680. Рассматривая процессы электронного транспорта в ФС II и ФС I в совокупности, можно заключить, что поглощение двух квантов света двумя молекулами хлорофилла приводит к переносу одного электрона с Н20 на С02, из которого синтезируются углеводы.
В ходе двухтактного (в ФС II и ФС I) фотоэлектронного преобразования возникают молекулярный кислород, НАДФН (восстановленный НАДФ), а также АТФ. Это единственная, хотя и сложная (многоступенчатая) световая реакция в процессе фотосинтеза. Дальнейшие аутотрофные биосинтетические реакции не нуждаются в свете. Они происходят в зеленых растениях за счет энергии, заключенной в электронах, принадлежащих НАДФН и АТФ.
Лучше других реакций фотосинтеза изучен процесс образования шестиуглеродной молекулы глюкозы из 6 молекул С02 и 6 молекул Н20 (реакция фиксации углекислого газа). 12 пар возбужденных электронов, необходимых для осуществления этой реакции, поступают от НАДФН. Дополнительным поставщиком свободной энергии служат 18 молекул АТФ. Вклад НАДФН и АТФ в фотосинтез одного моля глюкозы составляет соответственно 660 и 252 ккал.
Аналогично углеводам происходит фотосинтез всех биологически важных макромолекул. И здесь нужно подчеркнуть, что для созидания углеводов,белков, жиров, нуклеиновых кислот и других высокомолекулярных органических соединений из двуокиси углерода, воды, нитратов, сульфатов и прочих сравнительно простых веществ необходим колоссальный приток электронов к ним.
Углеводы, белки, жиры служат основными питательными веществами для гетеротрофов. В ходе катаболических процессов, также обеспечиваемых электрон-транспортными системами, освобождаются электроны примерно в таком же количестве, в каком они захватывались органическими веществами при их фотосинтезе. Электроны, освобождающиеся при катаболизме, переносятся на молекулярный кислород дыхательной цепью митохондрий. Здесь окисление сопряжено с фосфорилированием — синтезом АТФ посредством присоединения к АДФ остатка фосфорной кислоты (т. е. фосфорилирования АДФ). Этим обеспечивается энергоснабжение всех процессов жизнедеятельности
Лекция №6
Электромагнитные излучения и поля в природе, технике и жизни человека
На современном уровне развития науки известны два вида материи: вещество и поле. К веществу относятся формы материи, состоящие из элементарных частиц (протонов, электронов, нейтронов и др.), из которых состоят атомы и молекулы и, в конечном счете, все окружающие тела. Веществу свойственно наличие массы покоя. Поле — форма материи, посредством которой осуществляется взаимодействие между частицами вещества или телами. Полю свойственны непрерывность в пространстве и отсутствие массы покоя. Поле и вещество неразрывно связаны, образуя единую материю нашего мира.
Важнейшим универсальным свойством материи является корпускулярно-волновой дуализм. Всем микрообъектам присущи одновременно и корпускулярные, и волновые свойства. Впервые корпускулярно-волновой дуализм был установлен для света. Опыты по интерференции, дифракции света свидетельствовали о его волновой природе и подтверждали теорию Максвелла, установившую, что свет представляет собой электромагнитные волны.
Электромагнитной (ЭМ) волной называется процесс распространения в пространстве взаимоиндуцирующих друг друга переменных электрических и магнитных полей.
Основные положения об ЭМ поле были сформулированы английским физиком Максвеллом, объединившим в рамках единой теории электрические и магнитные явления. Им было показано, что ЭМ волны распространяются в среде с конечной скоростью, векторы напряженности электрического поля Е и магнитного поля H взаимно-перпендикулярны и фазы их колебаний одинаковы. Для плоской гармонической ЭМ волны, распространяющейся в изотропной среде без затухания:
Длиной волны называется расстояние между двумя ближайшими точками, колебания величин Е и Н в которых происходят в одинаковой фазе. Скорость распространения ЭМ волны в вакууме (с) есть величина постоянная.
Важнейшей характеристикой волны является интенсивность / — среднее количество энергии W , переносимое волной за единицу времени через единичную площадку, расположенную перпендикулярно направлению распространения волны.
Вместе с тем Планк показал, что для объяснения закона равновесного теплового излучения необходимо принять гипотезу о дискретном характере излучения, полагая, что энергия излучения кратна некоторой величине е, названная им квантом энергии.
В дальнейшем трудами Эйнштейна и ряда ученых было показано, что ЭМ-излучение не только испускается, но и распространяется квантами. Так возникло представление о частицах света фотонах, несущих квант энергии Е и движущихся со скоростью света.
Таким образом, было разрешено противоречие, возникшее еще в 17 столетии и содержащееся в утверждениях, что свет, с одной стороны, — это волна (Гюйгенс), а с другой - поток «корпускул» (Ньютон). ЭМ излучение обладает дуализмом, то есть одновременно и свойствами волны, и свойствами потока частиц. Корпускулярно-волновой дуализм присущ и другим микрочастицам, например электронам, нейтронам.
Один вид материи может превращаться в другой и обратно. Так известна реакция аннигиляции (уничтожения) двух античастиц: электрона и позитрона. В этой реакции вещество — электрон е- и позитрон е+ — превращаются в два кванта электромагнитного поля. В результате этой реакции могут образовываться два гамма-кванта, имеющие энергию не менее 0,51 МэВ каждый.
Известно и обратное превращение — реакция рождения пары. При этой реакции гамма-фотон высокой энергии (более 1,2 МэВ) при взаимодействии с электрическим полем ядра атома превращается в две частицы: электрон и позитрон. Таким образом, кванты электромагнитного поля превращаются в вещество. Реакции аннигиляции и рождения пар являются примером перехода одного вида материи в другой и обратно.
Понятие «физические поля окружающего мира», очевидно, является широким и может включать в себя многие явления в зависимости от целей и контекста рассмотрения. Если употреблять его в строго физическом смысле, то есть как вид материи, то следует иметь в виду, прежде всего электрическое, магнитное, электромагнитное, гравитационное поля и поле внутриядерных сил. В экологическом контексте в это понятие могут быть включены потоки ионизирующих частиц, акустические и вибрационные поля, атмосферные изменения и ряд других.
Вся биосфера Земли: простейшие, обширные царства растений и животных и человек — находится в окружении единого материального мира, составляющего ее среду обитания. Сфера обитания является неотъемлемым условием развития жизни и одновременно суммой факторов, влияющих на живые организмы и определяющих эволюцию живой природы. Одним из существенных факторов сферы обитания являются потоки излучений, действию которых подвергается все живое на Земле. Это электромагнитные волны, в безбрежном океане которых находится Земля, межзвездное и галактическое пространство, и ионизирующие излучения.
При авариях на АЭС или при ядерных взрывах в атмосферу могут выбрасываться различные радионуклиды. Эти изотопы могут накапливаться в организме, вызывая в нем нарушение деятельности, как отдельных органов, так и организма в целом.
Так, 131I накапливается в щитовидной железе, и уже 0,35 мг радиоактивного иода опасны для человека (при ежесуточной потребности около 150 мг). Изотоп 90Sr накапливается в костной ткани, а изотоп 137Cs равномерно распределяется в клетках организма.
Особую опасность представляют повышенные дозы радиоактивных излучений для кроветворной системы, пищеварительного тракта и желез внутренней секреции человека. Люди, работающие с излучением: в больницах, на АЭС, в лабораториях - могут получать дозу до 0,5 бэр в год.
Предельно допустимой биологической дозой для человека при профессиональном облучении считается 5 бэр в год. Минимальная летальная доза от «прямого у-луча» условно принята 600 бэр при облучении всего тела.
Ионизирующее излучение вызывает в биологических объектах различные патологические изменения, крайним проявлением которых является гибель облученных организмов. Степень и характер проявления радиобиологических эффектов зависят от различных радиационных факторов, характеризующих условия облучения, и от биологических факторов, характеризующих объект облучения.
К радиационным факторам относятся:
4. место расположения источника излучения по отношению к облучаемому организму;
5. вид ионизирующего излучения;
6. пространственное распределение дозы ионизирующего излучения в организме;
7. временное распределение дозы ионизирующего излучения;
8. доза облучения.
К биологическим факторам относятся: вид живого организма, возраст, пол, физиологическое состояние, используемая пища и т.д.
В зависимости от места расположения источника излучения по отношению к облучаемому биологическому объекту облучение может быть:
1) внешним — когда источник излучения находится во внешней среде вне облучаемого организма;
2) внутренним — когда облучение происходит в результате воздействия излучения от попавших в организм радиоактивных веществ (радионуклидов);
3) комбинированным — в реальной экологической обстановке встречается наиболее часто.
Проникновение (инкорпорирование) в организм радиоактивных веществ может осуществляться различными путями:
9. алиментарным, или пероральным путем (т.е. е продуктами питания и водой);
10. перкутанным путем (т.е. через кожные покровы);
11. через слизистые оболочки (глаза);
12. через раны.
Инкорпорированные в организм радионуклиды в зависимости от своих химических и физико-химических свойств могут распределяться в организме либо равномерно, либо неравномерно - органотропно, т.е. преимущественно в определенных органах и тканях. Для большинства радионуклидов характерна неравномерность распределения в организме, т.е. органотропность. Соответственно органы и ткани, в которых накапливается тот или иной радионуклид, подвергаются большему радиационному поражению при поступлении в организм этих радионуклидов.
Нужно отметить, что в некоторых случаях тип распределения радионуклида может меняться. В частности распределение кислорода, азота, водорода и углерода зависит от тех химических соединений, в составе которых они поступают в организм.
Снижение содержания попавших в организм радионуклидов происходит главным образом через желудочно-кишечный тракт (т.е. с калом) и почки (т.е. с мочой), в меньшей степени - через легкие (с выдыхаемым воздухом) и кожу (с потом), а также с молоком и плодом (например, яйца). Скорость выделения радионуклидов связана е их физико-химическими свойствами. Основное количество радионуклидов выводится в первые дни после поступления в организм. Быстрее всего выводятся из организма благородные газообразные радионуклиды, например радон (несколько часов). Из мягких тканей радионуклиды обычно выводятся быстрее, чем из костной ткани. Остеотропные радионуклиды наиболее долго удерживаются в организме.
Уменьшение количества радионуклидов в организме осуществляется по экспоненциальному закону в результате двух процессов: биологического выведения и радиоактивного распада.
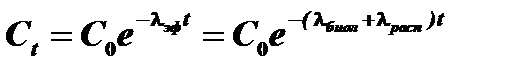
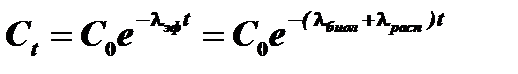
Константа 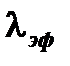 называется константой эффективного выведения и представляет собой сумму константы биологического выведения А и константы радиоактивного распада А . Вместо константы эффективного выведения часто пользуются величиной эффективного периода полувыведения. Эффективный период полувыведения представляет собой время, в течение которого содержание радионуклида в организме (или в органе) снижается вдвое. Константа эффективного выведения и эффективный период полувыведения связаны следующим соотношением:
называется константой эффективного выведения и представляет собой сумму константы биологического выведения А и константы радиоактивного распада А . Вместо константы эффективного выведения часто пользуются величиной эффективного периода полувыведения. Эффективный период полувыведения представляет собой время, в течение которого содержание радионуклида в организме (или в органе) снижается вдвое. Константа эффективного выведения и эффективный период полувыведения связаны следующим соотношением:
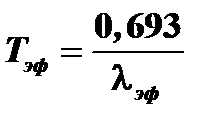
Эффективный период полувыведения существенно зависит от вида, возраста, функционального состояния организма.
Радиобиологические эффекты в значительной степени зависят от вида ионизирующего излучения, воздействующего на биологический объект. Наибольшее практическое значение имеют: рентгеновское излучение, гамма-излучение, бета-излучение, альфа-излучение, нейтронное излучение.
При внешнем облучении наибольшую опасность представляют те виды ионизирующих излучений, которые обладают наиболее высокой проникающей способностью, т.е. нейтронное, гамма- и рентгеновское излучения.
Альфа-излучение из-за низкой проникающей способности (в живой ткани - до 130 мкм) практически не представляет опасности при внешнем облучении организмов, размеры которых значительно превышают проникающей способности а-излучения. Наружный слой кожи, образованный отмершими клетками, ткань одежды практически полностью задерживают а-из лучение. Однако при внешнем облучении организмов, сравнимых по размерам с величиной проникающей способности a-излучения (например, одноклеточных организмов), а также при внутреннем облучении более крупных биологических объектов (т.е., когда процесс а-распада радионуклидов, попавших в организм, осуществляется в непосредственной близости от жизненно важных клеток организма) a-излучение является чрезвычайно опасным. В этих случаях a-излучение при одинаковой поглощенной дозе гораздо опаснее у-излучения - в среднем способность a-излучения повреждать клетки в 20 раз выше, чем у у-излучения.
Проникающая способность 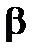 -излучения занимает промежуточное положение между а- и
-излучения занимает промежуточное положение между а- и 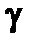 -излучениями: пробег в живой ткани составляет 1-1,5 см. Поэтому при внешнем облучении относительно крупных организмов (например, человека) поражающему действию
-излучениями: пробег в живой ткани составляет 1-1,5 см. Поэтому при внешнем облучении относительно крупных организмов (например, человека) поражающему действию 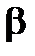 -излучения подвержены в основном только внешние ткани — кожа и глаза. Наибольшую опасность
-излучения подвержены в основном только внешние ткани — кожа и глаза. Наибольшую опасность 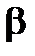 -излучение представляет (как и a-излучение) при внутреннем облучении. Способность
-излучение представляет (как и a-излучение) при внутреннем облучении. Способность 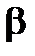 -излучения повреждать биологические клетки такая же, как у
-излучения повреждать биологические клетки такая же, как у 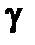 -излучения (при одинаковой поглощенной дозе).
-излучения (при одинаковой поглощенной дозе).
Пространственное и временное распределение дозы ионизирующего излучения в организме
Важным фактором, в значительной степени определяющим радиобиологический эффект, является пространственное распределение поглощенной дозы ионизирующего излучения в облучаемом организме. В зависимости от того, подвергается ли воздействию излучения весь организм или только какая-либо его часть, облучение разделяют на следующие типы:
13. общее (или тотальное) облучение — воздействию излучения подвергается все тело;
14. субтотальное облучение — воздействию излучения подвергается большая часть тела при защитном экранировании (например, свинцовыми пластинами) отдельных его областей или органов (например, головы, области живота, грудной клетки, конечностей, половых органов, и т.д.);
15. парциальное облучение — воздействию излучения подвергается отдельная область тела (например, голова, живот, грудная клетка и т.д.);
16. локальное облучение — воздействию узких пучков излучения подвергается отдельный орган или небольшой участок тела.
Все перечисленные типы облучения применяют в экспериментальной радиобиологии. В частности субтотальное, парциальное и локальное облучения в экспериментальной радиобиологии применяют для оценки значения нарушения функционирования отдельных органов и тканей в развитии лучевого поражения организма. При аварийных ситуациях происходит обычно либо общее, либо парциальное облучение. Локальное и парциальное облучения используют в лучевой терапии для лечения различных заболеваний, главным образом злокачественных опухолей.
Летальный исход для организма при общем облучении наблюдается обычно при более низких дозах, чем при других типах облучения. Поэтому локальное или парциальное облучение (особенно тех областей тела, которые не играют определяющего значения для выживания организма) даже в таких высоких дозах, которые значительно превышают смертельную дозу, характерную для общего облучения, могут не привести к летальному исходу.
Следует отметить, что общее облучение не подразумевает наличия равномерности облучения всего организма. Даже в условиях помещения облучаемого организма в равномерное поле глубокопроникающего 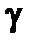 - или рентгеновского излучения более удаленные от источника излучения области тела получают меньшую поглощенную дозу излучения вследствие экранирования их менее удаленными областями тела. Степень неравномерности облучения в первом приближении можно оценить учитывая, что живая ткань на 90% состоит из воды, а слой половинного ослабления для наиболее часто используемых в радиобиологических экспериментах жестких рентгеновских лучей е энергией 250 кэВ составляет для воды 5,5 см. Для достижения большей равномерности облучения внутри крупных биологических объектов вместо рентгеновских лучей используют у-излучение ^Со (обладающее энергией 1,17 и 1,33 МэВ); слой половинного ослабления в воде в этом случае составляет несколько более 10 см. Большую равномерность облучения крупных биологических объектов достигают также путем использования двустороннего, четырехстороннего или многостороннего облучения. Принято считать, что облучение является равномерным, если различия в распределении поглощенной дозы в облучаемом организме не превышает ±10%. Равномерное облучение встречается практически исключительно только в экспериментальных условиях. В большинстве аварийных ситуаций наблюдается неравномерное облучение.
- или рентгеновского излучения более удаленные от источника излучения области тела получают меньшую поглощенную дозу излучения вследствие экранирования их менее удаленными областями тела. Степень неравномерности облучения в первом приближении можно оценить учитывая, что живая ткань на 90% состоит из воды, а слой половинного ослабления для наиболее часто используемых в радиобиологических экспериментах жестких рентгеновских лучей е энергией 250 кэВ составляет для воды 5,5 см. Для достижения большей равномерности облучения внутри крупных биологических объектов вместо рентгеновских лучей используют у-излучение ^Со (обладающее энергией 1,17 и 1,33 МэВ); слой половинного ослабления в воде в этом случае составляет несколько более 10 см. Большую равномерность облучения крупных биологических объектов достигают также путем использования двустороннего, четырехстороннего или многостороннего облучения. Принято считать, что облучение является равномерным, если различия в распределении поглощенной дозы в облучаемом организме не превышает ±10%. Равномерное облучение встречается практически исключительно только в экспериментальных условиях. В большинстве аварийных ситуаций наблюдается неравномерное облучение.
Для большинства радиобиологических эффектов исключительно важное значение имеет временное распределение поглощенной дозы, т.е. длительность облучения, а также наличие или отсутствие перерывов в облучении. Основным показателем, характеризующим распределение поглощенной дозы во времени, является мощность поглощенной дозы (часто говорят мощность дозы облучения). Мощность поглощенной дозы - это отношение приращения поглощенной дозы излучения dD за интервал времени dt к этому интервалу времени. Единица мощности поглощенной дозы D в системе СИ - грей в секунду (Гр/с). На практике единицу поглощенной дозы часто применяют с кратными или дольными приставками (обычно - кило, санти, милли и микро), а в качестве единицы времени используют также минуту, час, сутки, неделя, год. Поэтому в качестве внесистемных единиц мощности поглощенной дозы часто используют Гр/мин, еГр/еут, кГр/чае и т.п. В качестве единицы поглощенной дозы ранее использовали внесистемную единицу рад, а в качестве единицы мощности поглощенной дозы - рад/мин, рад/час, крад/год и т.п. (эти внесистемные единицы можно встретить иногда и в современной литературе).
В зависимости от длительности облучения, величины используемой мощности поглощенной дозы, наличия или отсутствия перерывов между облучениями условно можно выделить следующие 4 основные типа облучения:
17. 1) острое (или кратковремееное) облучение - лучевое воздействие при пролонгированное (или протяженное) облучение — лучевое воздействие при сравнительно небольшой мощности дозы (от 0,1 Гр/час и ниже) длительностью от 2 часов до нескольких недель;
18. дробное (или фракционированное) облучение - многократное лучевое воздействие с любой мощностью дозы (чаще при высокой мощности дозы в каждой фракции) с временными интервалами между фракциями облучения;
19. хроническое облучение — лучевое воздействие длительностью от нескольких месяцев до нескольких лет, осуществляемое либо постоянно (т.е. без перерывов) при низкой мощности дозы (порядка 0,01 Гр/сут и ниже), либо фракционирование (т.е. с некоторыми перерывами) в небольших разовых дозах при любой мощности дозы.
Разумеется, что эти 4 типа облучения не перекрывают всего многообразия возможных вариантов временного распределения поглощенной дозы и лишь условно обозначают наиболее характерные из них.
Многочисленные экспериментальные данные свидетельствуют, что фракционирование дозы и снижение мощности дозы (в диапазоне обычно используемых или встречающихся мощностей дозы) приводит обычно к ослаблению большинства биологических эффектов (при одной и той же поглощенной дозе), причем эффект может снижаться весьма существенно — в несколько раз. Наиболее сильное ослабление радиобиологических реакций наблюдается при снижении мощности дозы в диапазоне от нескольких единиц до долей рад/мин. В диапазоне от сотен до десятков рад/мин ослабление менее выражено. Однако, в области сверхвысоких мощностей дозы (выше нескольких тысяч рад/мин) возможна противоположная зависимость величины радиобиологического эффекта от мощности дозы, т.е. наблюдается снижение эффекта с увеличением мощности дозы. Таким образом, в целом зависимость величины радиобиологического эффекта от величины мощности дозы не является линейной, а имеет куполообразный характер.
Ослабление эффекта облучения при снижении мощности дозы или при фракционировании дозы связано с осуществлением в организме восстановительных (репарационных) процессов, в результате которых часть радиационных повреждений клеточных структур к моменту следующего взаимодействия с ними кванта излучения успевает репарироваться. Эта репарация, однако, не успевает произойти при более высокой мощности дозы, т.е., когда попадание квантов излучения в клеточную структуру происходит более часто. В результате этого чаще возникают невосстанавливаемые (нерепарируемые, нерепарабельные) повреждения клеточных структур (пример — двунитиевые повреждения ДНК). По этой же причине ослабление эффекта облучения при снижении мощности дозы выражено даже полностью отсутствовать при действии плотноионизирующих излучений, вызывающих значительно более тяжелые, невосстанавливаемые повреждения.
Эффект ослабления более характерен также для тех тканей организма, которые обладают высокой пролиферативной активностью и соответственно более высоким уровнем восстановительных (репарационных) процессов. Особенно отчетливо этот эффект наблюдается при лучевом поражении слизистой тонкой кишки.
При лучевом воздействии на ткани с низким уровнем пролиферативной активности и восстановительных процессов (например, на нервную ткань, печень, мышцы) феномен ослабления эффекта облуч
Дата добавления: 2017-02-13; просмотров: 2386;
Поиск по сайту
Узнать еще
- Адаптация растений и животных к световому режиму.
- Анатомическое строение стебля однодольных и двудольных растений. Строение стебля двудольных и голосеменных древесных растений. Структура древесины
- Ассимиляция углерода (фотосинтез)
- Ассортимент зеленых насаждений. Виды посадок. Нормативы по размещению зеленых насаждений.
- Ассортимент растений, рекомендуемый для создания санитарно-защитных зон и озеленения города
- Б) Адаптации растений к тепловому режиму
- Б) Экологические группы растений по отношению к свету
- Бактериальный фотосинтез

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
