Регуляция секреции ФСГ и ЛГ
Образование и освобождение обоих гормонов стимулируется гипоталамическим декапепти-дом - гонадотропин-рилизинг-гормоном, секреция которого происходит эпизодически, что в основном и определяет импульсный характер секреции ЛГ и ФСГ.
У женщин эстрогены и прогестерон по механизму обратной связи влияют на секрецию ЛГ и ФСГ как на гипоталамическом, так и на гипофизарном уровне.
У мужчин тестостерон и эстроген, образованный в клетках Лейдига и в процессе метаболизма тестостерона, блокируют по механизму обратной связи синтез и секрецию гонадолиберина и го-надотропных гормонов гипофиза. Кроме этого, клетками гранулёзы фолликулов и клетками Сертоли вырабатывается белок ингибин, который тормозит гипофизарную секрецию ФСГ.
T1/2 ФСГ составляет примерно 150 мин, а T1/2 ЛГ - 30 мин.
Механизм действия и эффекты ФСГ и ЛГ
Гонадотропные гормоны ЛГ и ФСГ связываются с рецепторами на мембранах своих клеток-мишеней в яичниках и яичках, в результате чего происходит активация аденилатциклазной системы. Образующийся цАМФ активирует протеинкиназу, которая фосфорилирует белки, опосредующие эффекты ЛГ и ФСГ.
У женщин лютеинизирующий гормон стимулирует образование прогестерона клетками жёлтого тела, у мужчин - синтез тестостерона интерстициальными клетками Лейдига. ФСГ ускоряет развитие фолликулов в яичниках и образование эстрогенов, а действуя на клетки Сертоли, запускает процесс сперматогенеза.
Б. МУЖСКИЕ ПОЛОВЫЕ ГОРМОНЫ
Мужские половые гормоны (рис. 11-39) вырабатываются в основном в мужских половых железах - в интерстициальных клетках Лейдига семенников (95%). Небольшое количество анд-рогенов образуется в коре надпочечников.
Синтез андрогенов
Путь биосинтеза андрогенов в яичках и коре надпочечников одинаков. Предшественником андрогенов, как и других стероидных гормонов, служит холестерол (рис. 11-40), который либо поступает из плазмы в составе ЛПНП, либо синтезируется в самих железах из ацетил-КоА.
Отщепление боковой цепи холестерола и образование прегненолона -скорость-лимити-рующая реакция. Однако, в отличие от аналогичной реакции, протекающей в надпочечниках, эта стадия стимулируется ЛГ (а не АКТГ). ЛГ, связываясь с рецептором плазматической мембраны клеток Лейдига, активирует адени-латциклазу, увеличивая тем самым внутриклеточную концентрацию цАМФ, что в конечном итоге вызывает активацию фермента, который расщепляет боковую цепь холестерола между
С-20 и С-22.
Тестостерон.Превращение прегненолона в тестостерон катализируется пятью микро-сомальными ферментами и может протекать двумя путями: через образование дегидроэпиан-дростерона или через образование прогестерона (что, по-видимому, преобладает в семенниках человека).
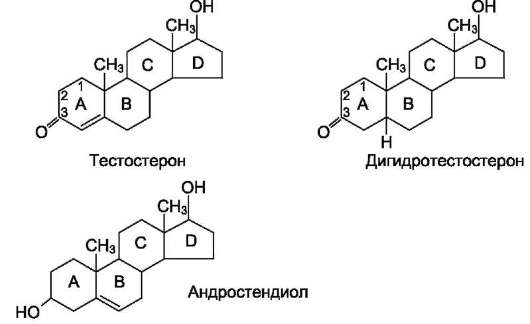
Рис. 11-39. Мужские половые гормоны.
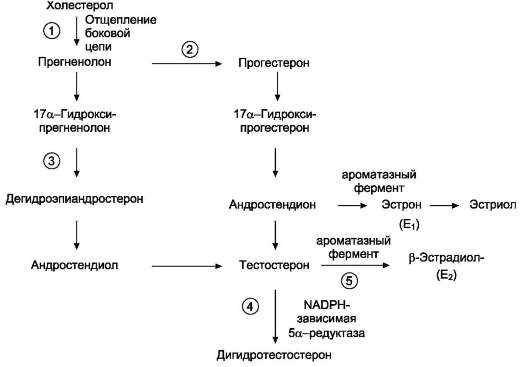
Рис. 11-40. Схема синтеза половых гормонов.Предшественником половых гормонов служит холестерол. Образование прегненолона происходит в результате отщепления боковой цепи холестерола (1). Превращение прегненолона в тестостерон может протекать двумя путями: через образование прогестерона (2) или дегидроэпиандростерона (3). Тестостерон служит предшественником дигидротестостерона (4). В некоторых периферических тканях небольшое количество тестостерона превращается в эстрадиол (5). В яичниках синтезируются женские половые гормоны, эстрогены и прогестины, среди которых наиболее активными являются 17β-эстрадиол и прогестерон. Ароматизация андрогенов протекает под действием ароматазного комплекса, содержащего цитохром Р450-оксидазу, и включает 3 реакции гидроксилирования с участием О2 и NADPH.
Суточная секреция тестостерона у мужчин составляет в норме примерно 5 мг и сохраняется на протяжении всей жизни организма. Гормон циркулирует в крови в связанном с белками плазмы состоянии: альбумином (40%) и специфически связывающим половые гормоны β-глобулином (называемым секс-гормонсвязывающим глобулином, СГСГ). Лишь 2% от общего количества гормона в крови транспортируется в свободном виде, и именно такие молекулы проявляют биологическую активность.
Дигидротестостерон.В семенных канальцах, предстательной железе, коже, наружных по-
ловых органах тестостерон служит предшественником более активного андрогена - ди-гидротестостерона (рис. 11-41, 11-42). Это превра-щение, в котором участвует примерно 4% тестостерона, происходит в результате восстановления двойной связи кольца А и 3-кето-группы при участии цитоплазматического фермента - NADPH-зависимой 5а-редуктазы. Семенники человека секретируют в сутки до 50-100 мкг дигидротестостерона. Однако большее количество гормона - следствие периферических превращений, и суммарная суточная секреция дигидротестостерона составляет
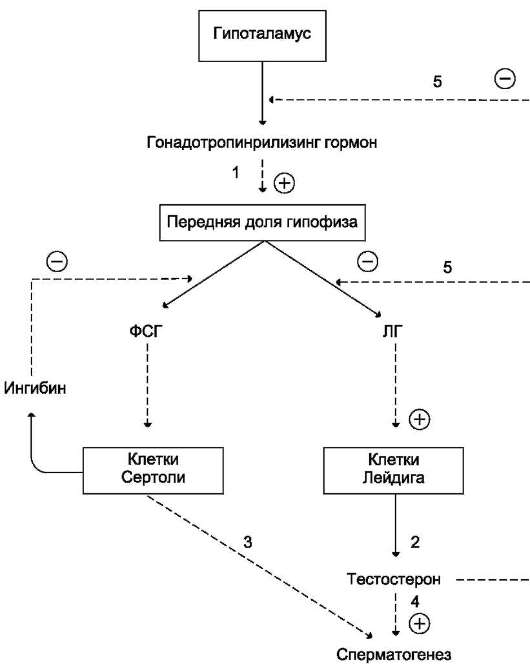
Рис. 11-41. Регуляция синтеза и секреции мужских половых гормонов.Синтез и секреция мужских половых гормонов регулируется гипоталамо-гипофизарной системой по механизму отрицательной обратной связи. Секреция ЛГ и ФСГ стимулируется гонадотропин-рилизинг гормоном. ЛГ ускоряет синтез и секрецию тестостерона клетками Лейдига, ФСГ стимулирует сперматогенез. Тестостерон стимулирует сперматогенез, ингибирует синтез и секрецию гонадотропин-рилизинг гормона и ЛГ.
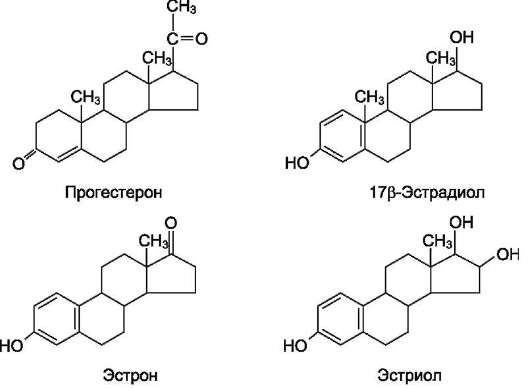
Рис. 11-42. Женские половые гормоны.
400 мкг, что почти в 10 раз меньше уровня секреции тестостерона.
В некоторых периферических тканях небольшое количество тестостерона превращается в эс-традиол. В качестве побочных продуктов клетки Лейдига также постоянно секретируют эстра-диол и прогестерон, хотя роль этих гормонов в развитии и поддержании функций размножения и формирования полового поведения у мужчин до настоящего времени не выяснена.
Дата добавления: 2017-01-08; просмотров: 4868;
Поиск по сайту
Узнать еще
- IV. Ликвидация гиповолемии и регуляция водно-солевого обмена
- Vi. регуляция водно-солевого обмена
- Vii. регуляция обмена ионов кальция и фосфатов
- АВТОНОМНАЯ САМОРЕГУЛЯЦИЯ ФЕРМЕНТАТИВНЫХ ПРОЦЕССОВ
- Аллостерическая регуляция каталитической активности ферментов
- Анаэробный гликолиз: понятие, этапы, общая схема, последовательность реакций, регуляция, энергетический баланс. Регуляция гликолиза
- Белковый обмен и его регуляция. Понятие об азотном балансе.
- Беременность, стадии беременности и ее регуляция

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
