Генетические процессы в малых популяциях
Советские ученые С.С. Четвериков и Т. Добржанский создали современную (синтетическую) теорию эволюции, которая объединила основные положения теории Ч. Дарвина и современные данные биологии, общей и популяционной генетики и теории информации. Кратко перечислим основные положения этой теории:
▪ элементарной единицей эволюции является популяция;
▪ мутации дают элементарный эволюционный материал;
▪ далее вступают в действие другие элементарные эволюционные факторы — дрейф генов, популяционные волны, изоляция и естественный отбор, изменяющие частоты генов в популяциях.
Эти процессы интенсивно протекают в малых популяциях. Они наблюдаются и в больших популяциях, однако при большой выборке очень мало влияют на изменение частот генов, поэтому в таких популяциях сохраняется действие закона Харди-Вайнберга.
Мутации изменяют частоту генов в популяциях. Частота естественного мутирования гена – 10-5—10-7 на поколение. Вследствие большого количества генов у человека (порядка 30 000) до 10% его гамет несут мутантные гены. Доминантные мутации проявляются уже в первом поколении и сразу же подвергаются действию естественного отбора. Рецессивные мутации (возникают несколько чаще) сначала должны накопиться в популяции и только с появлением рецессивных гомозигот начинают проявляться фенотипически и подвергаться действию естественного отбора. Насыщенность природных популяций рецессивными мутациями имеет большое значение для выживания вида (С.С. Четвериков, 1926). Например, с началом применения первых антибиотиков часть болезнетворных бактерий уже имела мутантные формы, нечувствительные к ним, благодаря чему они выжили в изменившихся условиях среды. Накопление мутантных аллелей способствует комбинативной изменчивости, приводящей к генетической гетерогенности (генетическому полиморфизму) природных популяций. Средняя степень гетерозиготности у растений составляет 17%, у беспозвоночных — 13,4%, у позвоночных — 6,6%, у человека — около 6,7%. Мутационный процесс обеспечивает разнообразие эволюционного материала.
Тип мутации определяет характер эволюции. Например, произошла мутация локуса рецессивного гена и наряду с генотипом аа появился генотип Аа: аа → Аа. Возможные варианты скрещивания:
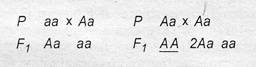
В результате в популяции появляются доминантные гомозиготы (в записи скрещивания они подчеркнуты).
Второй пример: произошла мутация локуса доминантного гена, и это привело к появлению гетерозиготной особи Аа: АА → Аа. Возможные варианты скрещивания:
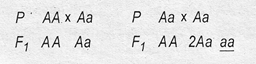
Такая мутация приводит к появлению рецессивных гомозигот (в записи скрещивания они подчеркнуты).
Насыщенность популяций рецессивными генами, снижающими приспособленность отдельных особей к условиям существования, которые элиминируются естественным отбором, называется генетическим грузом. Наличием генетического груза в человеческих популяциях объясняется появление до 5% потомков с генетическими дефектами.
Популяционные волны, или волны жизни (С.С.Четвериков, 1905), — это периодические колебания численности природных популяций в связи с колебаниями факторов внешней среды (смена времен года, урожайные и неурожайные годы и т. п.; рис. 8.).
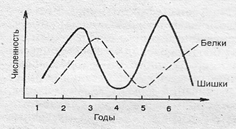
Рис. 8.1. Схема популяционных волн
Например, в неурожайные годы значительно уменьшается количество шишек на хвойных деревьях, вслед за чем происходит интенсивное снижение численности популяций белок. Популяционные волны являются результатом борьбы за существование, значительно усиливают естественный отбор и изменяют генетическую структуру популяций, элиминируя из нее менее приспособленные особи.
Дрейф генов (Н.П. Дубинин, Д.Д. Ромашов, С. Райт, Р. Фишер, 1931—1932) — это случайные колебания частот генов в малых популяциях. Предположим, что на необитаемый остров попало зерно гетерозиготного самоопыляемого растения. Исходная популяция будет состоять на 100% из гетерозиготных особей (Аа). В первом поколении будет уже только 50% гетерозиготных особей:
Р Aa x Аа 100% гетерозигот
F1 АА + 2Аа+аа 50% гетерозигот
F2 AA +AA +2Аа +аа+аа 25% гетерозигот
Гомозиготы (АА и aа) дадут только гомозиготных потомков, а гетерозиготы — расщепление 1:1 (поровну гомо- и гетерозигот), поэтому в F2 уже будет 25% гетерозигот. Таким образом, при самоопылении с каждым поколением количество гетерозигот уменьшается вдвое, что в конечном итоге приводит к гомозиготизации популяции. Если рецессивные (или доминантные) гомозиготы будут иметь преимущество в выживании, то со временем образуется чистая линия.
Иллюстрацией дрейфа генов может служить потеря фамилии в семьях, где рождается несколько девочек, которые, выходя замуж, берут фамилию мужа. Теоретически рождение мальчика или девочки равновероятно, но в малой выборке (семье) в силу случайных причин это равновесие может нарушаться.
Изоляция— это ограничение свободы скрещивания. Она способствует дивергенции — разделению популяций на отдельные группы и изменению частот генотипов. Различают географическую (горные хребты, реки, проливы и т. п.), генетическую (неполноценность гибридов, различные наборы хромосом), экологическую (различные экологические ниши, размножение при разных температурах) и морфофизиологическую (различия в строении половых органов) изоляцию.
В человеческих популяциях более существенными являются религиозные и морально-этические ограничения свободы браков. В малых человеческих популяциях (демах, изолятах) наблюдаются инбридинг (браки между генетическими родственниками) и дрейф генов. Родственные браки подразделяют на:
▪ инцестные (запретные) — браки между родственниками первой степени родства (родные брат и сестра, отец и дочь, мать и сын); они запрещены законодательствами многих стран и религиями;
▪ кровнородственные — браки между родственниками второй-третьей степени родства (двоюродные и троюродные братья и сестры). Эти браки нежелательны, они приводят к инбредной депрессии, так как у родственников высока степень вероятности гетерозиготности по одному и тому же рецессивному патологическому гену. Например, частота проявления фенилкетонурии при неродственных браках.составляет 1:15 000, а при родственных — 1:7000, альбинизма — 1:40 000 и 1:3000 соответственно.
Мерой генетических последствий инбридинга служит коэффициент инбридинга— вероятность того, что у какой-либо особи в данном локусе окажутся две аллели, идентичные по происхождению. У детей одной супружеской пары вероятность одинаковых аллелей в одном локусе равна 1/2. У их детей эта вероятность становится равной 1/4 (1/2x1/2). При вступлении в брак двоюродных сибсов (братьев и сестер) коэффициент инбридинга равен уже 1/16 (1/4x1/4).
В человеческих изолятах большая роль принадлежит так называемому "эффекту родоначальника", т. е. особенностям генотипов людей, основавших изолят. Если у основателей изолята имелись рецессивные летальные гены, то в условиях инбридинга они могут получить широкое распространение и привести изолят к вымиранию.
Изоляты известны даже в современных больших городах. В секте мормонов-менонитов в США, насчитывающей несколько тысяч человек выходцев из Голландии, высока частота генов карликовой хондродистрофии. В одной деревне в Швейцарии среди 2200 жителей имеется 50 глухонемых и 200 человек с генетически обусловленными дефектами слуха. В Южно-Африканской Республике среди белого населения часто встречается наследственное заболевание порфирия. Предполагают, что этим заболеванием страдала семья переселенцев из Голландии, прибывшая сюда в XVII в. Вследствие изоляции и дрейфа генов у североамериканских индейцев отсутствует ген III группы крови (1В) и преобладает 1(0) группа крови (ген 10). В одном из индейских племен концентрация гена группы крови П(А) составляет 80%. Частота встречаемости резус-отрицательных людей в Европе составляет 14%, а в Японии — 1%, что также является следствием изоляции и дрейфа генов.
Аутбридинг — неродственные браки, при которых поддерживается высокий уровень гетерозиготности. Повышению гетерозиготности человеческих популяций способствует миграция, масштабы которой огромны, особенно в последние десятилетия. Иммиграция поставляет новые аллели или новые комбинации генотипов, а эмиграция изменяет соотношение различных генотипов в популяции.
Естественный отбор элиминирует из популяции менее удачные комбинации генов и избирательно сохраняет более удачные генотипы, тем самым изменяя частоту генов в популяциях.
Для сохранения постоянства закона Харди-Вайнберга необходимо, чтобы каждая особь вносила свой вклад в генофонд будущих поколений. Интенсивность естественного отбора даже в современных человеческих популяциях довольно высока: спонтанные аборты составляют примерно 45% всех зачатий (30% — гибель зигот и 15% — гибель эмбрионов и плодов), мертворождения — 3%, ранняя детская смертность — 2%, не вступают в брак около 10% людей, примерно 10% браков бесплодны. Таким образом, окало 70% потенциальных инвесторов не вносят своего вклада в генофонд будущих поколений.
Различают 3 основные формы естественного отбора: движущий, стабилизирующий и дизруптивный.
Движущий отбор происходит при постепенном изменении факторов внешней среды или их колебаниях. Он устраняет неприспособленные формы и сохраняет отклонения, обеспечивающие адаптацию организмов к изменяющимся условиям; происходит смена нормы реакции или ее расширение (рис. 8.2). В качестве типичного примера можно привести постепенное вытеснение темноокрашенной формой бабочки березовой пяденицы исходной светлой формы, происходящее вследствие потемнения стволов берез от сажи и копоти.
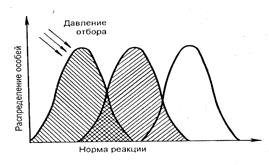
Рис. 8.2. Схема действия движущего отбора
Стабилизирующий отбор наблюдается при относительном постоянстве условий окружающей среды и направлен на сохранение в популяции среднего значения признака. Происходит сужение нормы реакции (рис. 8.3). Например, во время бури погибают преимущественно птицы с длинными и короткими крыльями, а выживают особи с крыльями средних размеров.
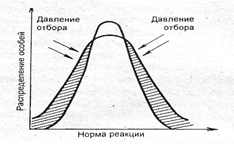
Рис. 8.3. Схема действия стабилизирующего отбора
Дизруптивный (разрывающий) отбор протекает на фоне резкого изменения условий существования. Он направлен против среднего значения признака и благоприятствует двум или нескольким направлениям изменчивости; при этом значительно изменяется норма реакции (рис. 8.4). Типичный пример — сохранение на открытых океанических островах либо бескрылых насекомых, либо насекомых с мощными крыльями, позволяющими противостоять порывам ветра.

Рис. 8.4. Схема действия дизруптивного отбора
Помимо естественного отбора в популяциях (в том числе и человеческих) может действовать и контротбор — отбор признаков, неблагоприятных в обычных условиях среды. Например, в странах Западной Африки частота патологического гена серповидно-клеточной анемии довольно высока, в то время как в странах умеренного климата он не встречается. Такая распространенность данного гена объясняется устойчивостью гетерозигот к тропической малярии. Гомозиготы по гену НbА (НbАНbА) имеют нормальный гемоглобин А, хорошо переносящий кислород, но они болеют и погибают от тропической малярии. Гомозиготы по гену НbS(HbSHbS) болеют серповидно-клеточной анемией, их гемоглобин плохо связывает и переносит кислород и они погибают в раннем детском возрасте от его недостатка. Гетерозиготы (НbА НbS) содержат и гемоглобин А (хорошо переносит кислород), и гемоглобин S (обеспечивает устойчивость к тропической малярии), поэтому выживают в эндемичных по тропической малярии районах.
Раздел II
ОСНОВЫ
Дата добавления: 2016-11-26; просмотров: 6048;
Поиск по сайту
Узнать еще
- I.1.3 ТЕРМОДИНАМИЧЕСКИЙ ПРОЦЕСС. ИЗОПРОЦЕССЫ
- V. Новообразования (доброкачественные и злокачественные) и диспласти-ческие (опухолевые) процессы.
- XIX. ХОЛОДИЛЬНЫЕ ПРОЦЕССЫ
- Автогенетические процессы и половой отбор
- АДИАБАТИЧЕСКИЕ ПРОЦЕССЫ В АТМОСФЕРЕ
- Баромембранные процессы.
- Бизнес-процессы. Классификация.
- Биогеохимические циклы (биогеохимические круговороты) -циклические процессы обмена веществ между различными компонентами биосферы, обусловленные жизнедеятельностью организмов.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
