Биогеохимические циклы (биогеохимические круговороты) -циклические процессы обмена веществ между различными компонентами биосферы, обусловленные жизнедеятельностью организмов.
Термин "биогеохимия» предложен русским ученым В.И. Вернадским и означает область науки об обмене веществ между живым и неживым веществом биосферы («био» относится к живым организмам, а "гео" - к горным породам, воздуху и воде). Геохимия изучает химический состав Земли и миграцию элементов между различными частями биосферы: литосферой, гидросферой и атмосферой.
Для нормального существования большинства экосистем и организмов, их населяющих, максимальное значение имеют круговороты таких элементов, как водород, углерод, азот, сера и фосфор, входящих в состав любого живого вещества.
В круговоротах любых химических элементов и веществ различают две части или два «фонда»:
1) резервный фонд - большая масса медленно движущихся в биогеохимическом цикле веществ;
2) обменный (подвижный) фонд - меньшая, но более активная масса вещества, для которого характерен быстрый обмен между живыми организмами и их непосредственным окружением.
В целом биогеохимические циклы обычно подразделяют на два основных типа:
1) круговорот газообразных веществ с резервным фондом в атмосфере или гидросфере (океане); 2)осадочный цикл с резервным фондом и земной коре. Резервные фонды в атмосфере и гидросфере легко доступны, поэтому такие круговороты относительно устойчивы. Осадочные биогеохимические циклы, как правило, менее стабильны.
Удивительное постоянство процентного содержания различных химических элементов в компонентах экосистемы исторически обусловлено существованием непрерывных и сбалансированных круговоротов веществ, что создает возможность для саморегуляции (гомостаза) системы и поддержания ее устойчивости.
Процессы новообразования органического вещества в ходе фотосинтеза и процессы его разрушения (распада) определяют скорость и сбалансированность круговоротов элементов в биосфере и происходят только за счет поступающей извне солнечной энергии. Следовательно, скорость и направление циклического движения элементов в экосистеме определяются потоками энергии, проходящей через биологическое сообщество.
Обобщенная схема биогеохимических циклов в сочетании с упрощенной схемой потока энергии (рис. 10.1) показывает, как однонаправленный поток энергии приводит в движение круговорот вещества. Обращает на себя внимание тот факт, что химические элементы, вовлеченные в процесс круговорота, многократно проходят один и тот же путь, а энергия течет лишь в одну сторону.
На рис. 10.1 резервный фонд обозначен как фонд элементов питания, а обменный фонд представлен темным кольцом, идущим от автотрофов к гетеротрофам и от них снова к автотрофам. Иногда резервный фонд называют недоступным, а активный обменный фонд - доступным. Например, агрономы обычно измеряют плодородие почвы, оценивая концентрацию в почве тех форм элементов питания, которые непосредственно доступны для растений.
Обменный фонд образуется за счет веществ, которые возвращаются в круговорот двумя основными путями - либо в результате прижизненных выделений во внешнюю среду продуктов метаболизма животными и растениями, либо при разрушении (минерализации) мертвого органического вещества (детрита) микроорганизмами.
Влияние человека на биогеохимические круговороты заключается в том, что при антропогенном вмешательстве эти процессы могут перестать быть замкнутыми и в одних местах боиосферы может возникнуть недостаток, а в других - избыток каких-либо веществ. В конечном счете меры по охране природных ресурсов должны быть направлены на предотвращение нарушений цикличности, т.e. сбалансированности круговоротов важнейших элементов в биосфере. Знание особенностей биогехимических циклов - необходимое условие рационального использования природных ресурсов и сохранения природных экосистем.
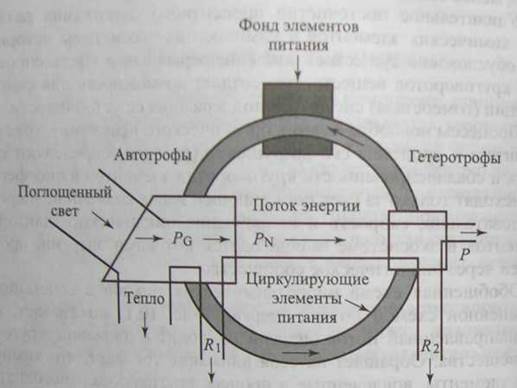
Рис. 10.1. Схема биогеохимического цикла на фоне упрощенной схемы потока энергии:
РG- валовая первичная продукция; РN- чистая первичная продукция (может быть потреблена гетерофами в самой системе или же экспортирована); Р - вторичная продукция, R1 -дыхание автотрофов (растений); R2 - дыхание гетеротрофов (животных и бактерий)
Любую экосистему можно представить в виде ряда блоков, через которые проходят различные вещества. В круговоротах минеральных веществ в экосистеме, как правило, участвуют три активных блока: живые организмы, мертвый органический детрит, доступные неорганические вещества в среде обитания.
Рассмотрим биогеохимический циклы азота, фосфора и серы. Биогеохимический цикл азота (биогенного элемента, входящего в состав белков и нуклеиновых кислот) может служить примером очень сложного хорошо сбалансированного цикла газообразного вещества. Биогеохимический цикл фосфора - осадочный цикл с менее совершенной регуляцией круговорота фосфора.
Биогеохимический круговорот серы служит примером функциональной связи между атмосферой, водой и земной корой, так как сера активно циркулирует в каждом из этих «резервуаров» и между ними. В круговoротax азота и серы ключевую роль играют микроорганизмы.
Круговорот азота, включающий как газовую, так и минеральную фазу, несмотря на большое число участвующих в нем организмов, обеспечивает быструю циркуляцию азота в различных экосистемах (рис. 10.2).
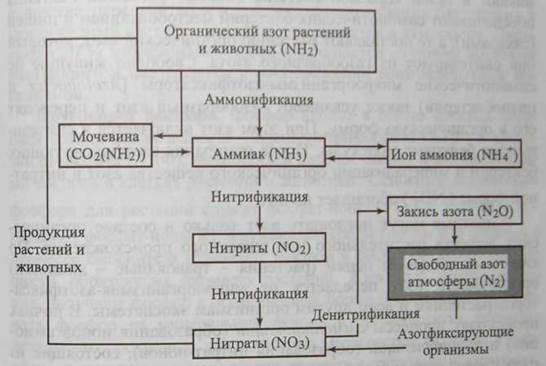
Рис. 10.2. Схема круговорота азота (серый прямоугольник – резервный фонд азота)
Основной источник и резервуар азота - атмосфера, масса которой на 79% состоит из этого элемента. Участие живых организмов в круговороте азота подчинено строгой иерархии: только определенные виды микроорганизмов (бактерий) осуществляют биохимические процессы трансформации соединении азота на отдельных ключевых этапах этого цикла.
Большинство организмов, обитающих в биосфере, непосредственно не может использовать газообразный молекулярный азот (N2). Растения усваивают азот только в составе нитрат - ионов (NО3-) или ионов аммония (NH4+). Нитраты образуются в основном в результате жизнедеятельности микроорганизмов - азотфиксаторов, к которым относятся симбиотические бактерии рода Rhizobium, живущие в клубеньках на корнях бобовых растений, бактерии рода Azotobacter, обитающие в почве; и цианобактерии (сине-зеленые). Все микроорганизмы - азотофиксаторы способны фиксировать атмосферный азот благодаря очень сложному обмену веществ, включающему в качестве катализаторов молибден и гемоглобин. Симбиотические микроорганизмы-азотофиксаторы проникают в ткани корневой системы бобовых растений. Растения обеспечивают симбиотических бактерий местообитанием и пищей (сахарами), а те поставляют растению органический азот, который они синтезируют из газообразного азота. Свободно живущие не симбиотические микроорганизмы - азотофиксаторы (Azotobacter в цианобактерии) также усваивают газообразный азот и переводят его в органическую форму. При этом азот включается в синтезируемые белковые молекулы. После отмирания азотофиксирующих бактерий и минерализации органического вещества азот в нитратной форме (NO3-) обогащает почву.
Животные могут поглощать азот только в составе органических веществ растительного или животного происхождения. По типичным пищевым цепям (растения - травоядные - хищники) органический азот передается от микроорганизмов - азотофиксаторов растениям и всем другим организмам экосистемы. В почвах происходят процессы аммонификации (образования ионов аммония) и нитрификации (образования нитрат - ионов), состоящие из ряда последовательных реакций, в ходе которых при участии разных групп микроорганизмов происходит разрушение мертвого органического вещества.
Молекулярный азот возвращается в атмосферу и биогеохимический цикл азота замыкается в процессе жизнедеятельности бактерий - денитрификаторов рода Pseudomonas, восстанавливающих нитраты до свободного азота и кислорода в бескислородных (анаэробных) условиях.
Нитраты постоянно образуются из молекулярного азота в небольших количествах без участия микроорганизмов-азотфиксаторов при электрических грозовых разрядах в атмосфере. Затем эти нитраты выпадают с дождями на поверхность почвы. Еще одним источником поступления атмосферного азота в биогеохимический цикл -вулканы, компенсирующие потери азота, выключенного из круговорота при осаждении его на дно океанов.
Для того чтобы сопоставить масштабы различных процессов поступления атмосферного азота в биогеохимический цикл, необходимо иметь в виду следующее : среднегодовое поступление нитратного азота абиотического происхождения (грозовые разряды) из атмосферы в почву не превышает 10 кг/га, свободные микроорганизмы- азотофиксаторы вносят до 25 кг/га, в то время как симбиотические азотофиксирующие бактерии Rhizobium в среднем продуцируют до 200 кг/га.
Преобладающая часть азота, содержащегося в органическом веществе, перерабатывается денитрифицирующими бактериями в газообразный азот (N2) и вновь возвращается в атмосферу. Лишь около 10% минерального азота поглощается из почвы высшими растениями и оказывается в распоряжении многоклеточных организмов.
Круговорот фосфора. Фосфор входит в состав богатых энергией органических веществ - аденознитрифосфата (АТФ) и аденозиндифосфата (АДФ), являющихся переносчиками и аккумуляторами энергии в клетках растений и животных. Основным источником фосфора для растений служат фосфат-ионы (РО4-). Растения поглощают фосфат-ионы из окружающей среды (почвы или воды) и в процессе биосинтеза включают фосфор в состав оpганических веществ, образующих биомассу растений. Животные, поедая растения, получают фосфор в органической форме. Таким образом, переводя фосфор из минеральной формы в органическую, растения делают его доступным для животных. Круговорот фосфора в биосфере связан с процессами обмена веществ в растениях и животных. Этот важный биогенный элемент, содержание которого наземных частях растений и водорослях варьирует от 0.01 до 0,1%, а у животных от 0.1 % до нескольких процентов, в процессе циркуляции постепенно переходит из органических соединений в фосфаты, которые снова могут использоваться растениями (рис 10.3).
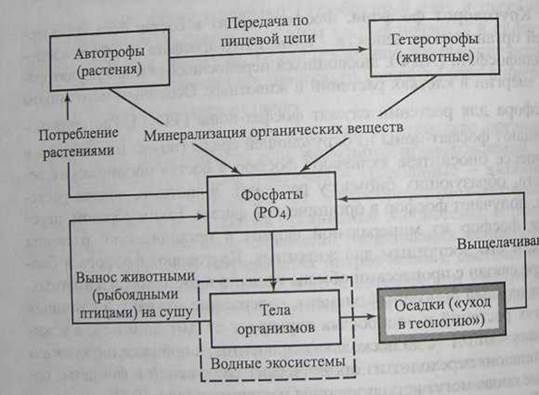
Рис. 10.3. Схема круговорота фосфора (серый прямоугольник - резервный фонд фосфора)
Если сравнить содержание фосфора в живом и неживом веществе биосферы, то окажется что диспропорция очень велика. Поэтому фосфор относится к числу наиболее дефицитных биогенных макроэлементов, определяющих развитие жизни.
Естественный биогеохимический круговорот фосфора в биосфере не сбалансирован. Основные запасы фосфора содержатся в горных породах (апатиты, фосфориты), из которых в процессе выщелачивания водорастворимые фосфаты (РО43-) попадают в наземные и водные экосистемы. Попадая в экосистемы суши, фосфор поглощается растениями из водного раствора в виде неорганического фосфат - иона (РО43-) и включается в состав различных фосфорорганических соединений. По пищевым цепям фосфорсодержащее органическое вещество переходит от растений к другим организмам экосистемы. Химически связанный фосфор попадает с остатками растений и животных в почву, где подвергается воздействию микроорганизмов и превращается в минеральные соединения фосфора, доступные растениям в ходе фотосинтеза. Вынос фосфатов из наземных экосистем в континентальные водоемы обогащает последние фосфором. Речной сток ежегодно выносит в Мировой океан около 2 млн. т фосфора.
В морских экосистемах минеральный фосфор переходит в состав фитопланктона, служащего пищей другим организмам моря, и накапливается в тканях морских животных, например, рыб. Часть органических соединений фосфора мигрирует по пищевым цепям в пределах небольших глубин, другая часть опускается на большие глубины в процессе осаждения мертвого органического вещества. Отмершие остатки организмов приводят к накоплению фосфора на разных глубинах. Отсюда следует, что фосфор, попадая в водоемы тем или иным путем, насыщает, а нередко и перенасыщает их экосистемы. Обратное движение фосфора из Мирового океана на сушу и в наземные водоемы ограничено (вылов рыб и других организмов человеком) и не компенсирует вынос фосфора с суши. И только в значительных временных интервалах, когда в процессе тектонического движения земной коры дно океанов становится сушей, происходит замыкание этого биогеохимического цикла.
Круговорот серы. Существуют многочисленные газообразные соединения серы, например сероводород (H2S) и сернистый ангидрид (SO3).
Однако преобладающая часть круговорота этого элемента имеет осадочную природу и происходит в почве и воде.
Подробная схема круговорота серы приведена на рис. 10.4.
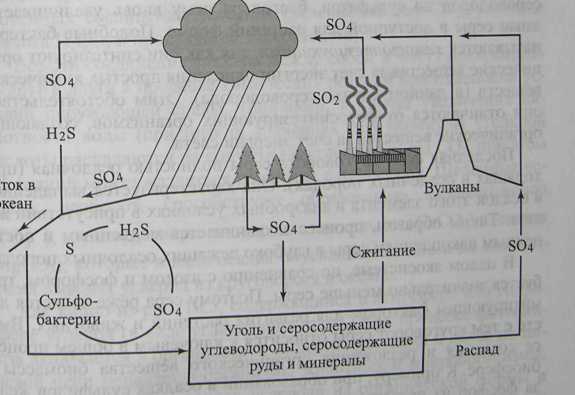
Рис 10.4. Схема круговорота серы
Основной источник серы, доступный живым организмам, - сульфаты (SO42- ). Многие сульфаты растворимы в воде, и это определяет доступность неорганической серы для растений, так как многие элементы (в том числе и сера) могут поступать в живые организмы только в растворенном виде. Растения, поглощая сульфаты, восстанавливают их и вырабатывают незаменимые серосодержащие аминокислоты (метионин, цистеин, цистин), играющие важную роль в создании третичной (пространственной) структуры белков. Животные и микроорганизмы, потребляя растительную биомассу в пищу, усваивают серосодержащие органические соединения.
При разложении мертвого органического вещества (опавшая листва, погибшие организмы, продукты выделения) гетеротрофными бактериями сера вновь переходит в неорганическую форму (преимущественно в виде сероводорода H2S). Некоторые бактерии могут вырабатывать сероводород из сульфатов в анаэробных (бескислородных) условиях. Другая немногочисленная группа бактерий может восстанавливать сероводород до элементарной серы (S).
С другой стороны, существуют бактерии, опять окисляющие сероводород до сульфатов, благодаря чему вновь увеличивается запас серы в доступной для растений форме. Подобные бактерии называются хемосинтезирующими, так как они синтезируют органические вещества за счет энергии окисления простых химических веществ (в данном случае сероводорода). Этим обстоятельством они отличаются от фотосинтезирующих организмов, создающих органические вещества за счет энергии света.
Последняя фаза круговорота серы полностью осадочная (проходящая в осадочных породах). Она характеризуется выпадением в осадок этого элемента в анаэробных условиях в присутствии железа. Таким образом, процесс заканчивается медленным и постепенным накоплением серы в глубоко лежащих осадочных породах.
В целом экосистеме, по сравнению с азотом и фосфором, требуется значительно меньше серы. Поэтому сера реже является лимитирующим фактором для развития растений и животных. Вместе с тем круговорот серы относится к ключевым в общем процессе создания разложения органического вещества биомассы в биосфере. К примеру, при образовании в осадках сульфидов железа фосфор из нерастворимой формы переходит в растворимую и становится доступным для фотосинтезирующих организмов. Это служит наглядным подтверждением того, что один круговорот связан с другим и регулируется им.
Круговорот углерода. Углерод в качестве важнейшего структурного элемента входит в состав любого органического вещества, поэтому его круговорот во многом определяет интенсивность образования и разрушения органического вещества в различных частях биосферы. В природе углерод существует в двух наиболее распространенных минеральных формах - в виде карбонатов (известняков) и в виде подвижной формы углекислого газа (yглекислоты, СО2). В биохимическом круговороте углерода атмосферный фонд углекислого газa относительно невелик (711 млрд. т) в сравнении с запасами углерода в океанах (39000 млрд. т), в ископаемом топливе (12000 млрд. т) и наземных экосистемах (3100 млрд. т).
Приблизительно 93 % углекислого газа находится в океане, который способен удерживать намного больше этого химического соединения, чем другие резервуары. Большая часть углекислоты, поступающей из атмосферы в поверхностные слои морской воды, взаимодействует с водой с образованием угольной кислоты и продуктов ее диссоциации. Таким образом, в океане постоянно существует карбонатная система - сумма всех неорганических растворенных соединений углерода (углекислый газ СО2, угольная кислота Н2СО3 и продукты ее диссоциации).
Все эти соединения связаны между собой и могут превращаться друг в друга в процессе химических реакций при изменении условий окружающей среды. Например, в случае увеличения кислотности воды (при низких значениях рН) молекулы угольной кислоты распадаются на воду Н2О и углекислый газ СО2, при этом последний может удаляться из океана в атмосферу. Щелочные условия, наоборот, способствуют образованию карбонат -ионов (СОз2-), труднорастворимых карбонатов кальция (СаСО3) и магния (MgCO3), которые в виде осадка опускаются на дно и на какое-то время выводят углерод из круговорота в океане.
Как видно из рис. 10.5, содержащийся в атмосфере или гидросфере углерод (в виде углекислого газа СО2) в процессе фотосинтеза включается в органическое вещество растений и далее по пищевой цепи попадает в организмы животных и микроорганизмы. Обратный процесс перехода углерода из органической формы в минеральную происходит во время дыхания всех организмов животных, и растений (окисление органического вещества до углекислого газа (СО2) и воды (Н2О)). Процесс высвобождения углекислого газа из органического вещества происходит не сразу, а постепенно, частями на каждом трофическом уровне. В почве очень часто биогеохимический цикл углерода замедляется, так как органические вещества минерализуются не полностью, а трансформируются в органические комплексы - гумус.
Особенность функционирования наземных экосистем - значительное и относительно долговременное накопление органической формы углерода в биомассе растений и животных, а также в гумусе. Таким образом, биомасса наземных экосистем также может рассматриваться как значительный запас углерода в биосфере.
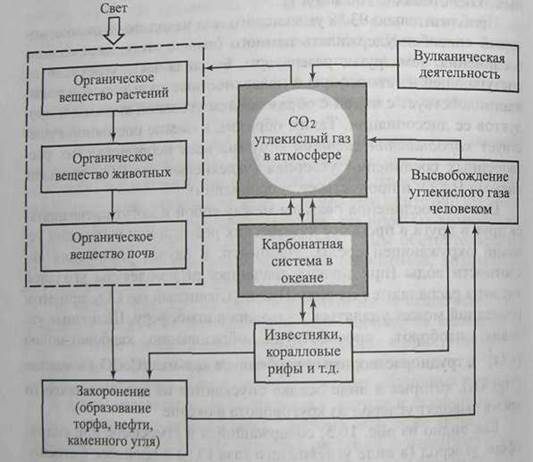
Рис. 10.5. Схема круговорота углерода (серые прямоугольники - резервные фонды углерода)
Океаническая ветвь биогеохимического цикла углерода имеет свои особенности, которые, учитывая значительный объем содержащегося в воде углерода, определяют важную роль Мирового океана в круговороте данного элемента. В водной среде в отличие от наземных экосистем основными фотосинтезирующими организмами являются одноклеточные микроскопические водоросли, парящие в водной толще (фитопланктон).
Жизнедеятельность организмов фитопланктона достаточно активна и сопровождается как накоплением органического углерода в виде биомассы, так и выделением растворенного органического углерода. Животные и бактерии потребляют эти органические формы углерода.
Особенностью функционирования водной экосистемы является быстрый переход органических форм углерода по пищевой цепи от одних организмов к другим. В отличие от наземных экосистем в океане не образуются значительные запасы органического углерода в биомассе живых организмов. Большая часть органического углерода в гидросфере вновь потребляется и в конце концов окисляется до минеральной формы - углекислого газа (СО2). Другая часть мертвого органического вещества (детрит) под действием силы тяжести оседает в глубокие слои водной толщи и откладывается на дне, где может долгое время сохраняться в виде органических осадков.
Небольшая часть органического вещества и содержащегося в нем углерода, по терминологии В.И. Вернадского, ускользает от круговорота и «уходит в геологию» - в отложения в виде торфа, угля, нефти и известняка в водных экосистемах.
Современный баланс углекислого газа в атмосфере представлен в табл. 10.1.
Таблица 10.1
Ежегодный баланс СО2 в атмосфере
| Составляющие баланса | Ежегодные поступления/убывания СО2 |
| Поступление в атмосферу: | |
| индустриальные выбросы в процессе вырубки лесов при эрозии почв | 6,41 1,08 0,91 |
| Поглощение из атмосферы: | |
| экосистемами суши (фотосинтез) Мировым океаном («карбонатная система») СО2, остающийся в итоге ежегодно в атмосфере | 4,05 1,05 3,3 |
Источник: Тарко А.М. Устойчивость биосферных процессов и принципов Ле Шателье // Доклады РАН. 1995. Т. 343. № 3. С. 123.
Таким образом, около из 6,41 млрд. т углекислого газа, ежегодно выбрасываемых промышленностью, 3,3 млрд. т, т.е. более 50% остается и атмосфере. За последние 150 лет это уже привело к увеличению содержания углекислоты в атмосфере более чем на 25% и вызвало стимуляцию парникового эффекта. В свою очередь изменение климатического режима Земли может привести и уже приводит к глобальному изменению климата.
В целом в биосфере в постоянном круговороте находится около 0,2% мобильного запаса углерода. Углерод биомассы обновляется за 12 лет, атмосферы - за 8 лет, что подтверждает высочайшую сбалансированность биогеохимического цикла углерода.
Контрольные вопросы и задания.
1. Что называется биогеохимическими циклами и как они связаны с экосистемами?
2. Охарактеризуйте резервный и обменный фонд в круговороте химических элементов.
3. Укажите блоки экосистем, через которые проходят биогеохимические циклы элементов.
4. В круговороте каких биогенных элементов ключевая роль принадлежит микроорганизмам?
5. Для каких элементов атмосфера является резервным фондом?
ЭКОЛОГИЯ ПОПУЛЯЦИЙ
Каждый биологический вид, существующий в природе, - это сложный комплекс внутривидовых групп организмов с однотипными чертами строения, физиологией и образом жизни. Такими внутривидовыми группами организмов являются популяции.
Популяция - группа организмов одного вида, способная поддерживать свою численность длительное время, занимающая определенное пространство и функционирующая как часть биотического сообщества экосистемы
Биотическое сообщество представляет собой совокупность популяций организмов разных видов, функционирующих как целостная система в определенном физико-географическом пространстве среды обитания.
Приспособительные возможности у популяции значительно выше, чем у слагающих ее индивидов. Популяция как биологическая единица обладает определенной структурой и функциями.
Популяция обладает биологическими свойствами, присущими как популяции в целом, так и составляющим ее организмам, и групповыми свойствами, проявляющимися только в целой группе. К биологическим свойствам популяции относятся, в частности, рост и участие в круговороте веществ. В отличие от биологических, групповые свойства: рождаемость, смертность, возрастная структура, распределение в пространстве, генетическая приспособленность и репродуктивная непрерывность (т.е. вероятность оставления потомков на протяжении длительного периода времени) - могут характеризовать только популяцию в целом.
Ниже представлены основные показатели популяции.
Плотность популяции - это численность популяции, отнесенная к единице пространства. Ее обычно измеряют и выражают числом организмов (численность популяции) или суммарной биомассой организмов (биомассой популяции) на единицу площади или объема, например, 500 деревьев на 1 га, 5 млн. микроводорослей на 1 м3 воды или 200 кг рыбы на 1га поверхности водоема.
Иногда бывает важно различить удельную, или экологическую плотность (численность или биомассу на единицу обитаемого пространства, т. е. фактически доступного для организмов конкретной популяции) и среднюю плотность (величину популяции, отнесенную к единице пространства в географических пределах обитания популяции). Например, средняя плотность лесных лягушек - это их численность, отнесенная к площади лесного массива. Однако эти животные обитают только в заболоченных участках леса, площади которых учитываются при расчете удельной плотности популяции.
Плотность популяции не является постоянной величиной - она изменяется с течением времени в зависимости от условий обитания, сезона года и т. д. Распределение организмов в пространстве, занимаемом популяцией, может быть случайным, равномерным и групповым. Чаще всего в природе встречаются различного рода скопления организмов одного вида (групповое распределение: семейные группы и стаи у животных, групповые заросли у растений).
Наиболее полное представление о плотности популяции дает комплексное использование показателей: численность особей хорошо характеризует их среднюю удаленность друг от друга; биомасса - концентрацию живого вещества; калорийность - количество связанной в организмах энергии. Как правило, плотность популяции растений выше, чем плотность популяции травоядных животных на той же территории. Чем крупнее организмы, тем больше их биомасса.
Плотность - одно из важнейших свойств популяции. От плотности популяции зависят дыхание, питание, размножение и многие другие функции отдельных организмов популяции. Чрезмерная плотность популяции ухудшает условия ее существования, снижая обеспеченность организмов пищей, водой, жизненным пространством и т. д. Отрицательно влияет на существование популяции и недостаточная ее плотность, которая затрудняет выбор особей противоположного пола, защиту популяции от хищников и т.д. (см. подробнее о массовых и групповых эффектах в лекции 6).
Существует ряд механизмов поддержания плотности популяций на нужном уровне. Главный из них - саморегуляция численности популяции по принципу обратной связи с количество и ограниченных жизненных ресурсов, в частности, пищи. Так, когда пищи становится меньше, рост особей замедляется, смертность возрастает, половая зрелость (т. е. способность к размножению) наступает позже, и в результате численность популяции и ее плотность снижается. Улучшение условий существования сопровождается я изменениями противоположного характера, и плотность популяции возрастает до определенного предела. Численность популяции может колебаться вследствие миграции, смены поколений, появления новых особей (благодаря рождению и вселению из других популяций) или в результате гибели. Изучение динамики численности популяции весьма важно для предсказания вспышек численности организмов вредителей или промысловых животных.
Численность популяции определяется в основном двумя противоположными явлениями - рождаемостью и смертностью.
Рождаемость - это способность популяции к увеличению численности. Она характеризует появление на свет новых организмов в процессе: рождения у животных, прорастания семян у растений, образования новых клеток в результате деления у микроорганизмов. Общее число новых молодых особей ( 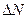 ), появившихся в популяции за единицу времени (Δt), называют абсолютной (общей) рождаемостью. Для сравнения рождаемости разных популяций используется понятие удельной рождаемости (b), выраженной числом новых особей на одну особь в единицу времени:
), появившихся в популяции за единицу времени (Δt), называют абсолютной (общей) рождаемостью. Для сравнения рождаемости разных популяций используется понятие удельной рождаемости (b), выраженной числом новых особей на одну особь в единицу времени:
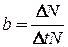
Так, для популяций человека в качестве показателя удельной рождаемости используют количество новорожденных детей, родившихся за 1 год на 1 тыс. населения.
Максимальная (потенциальная) рождаемость - это теоретический максимум скорости появления новых особей в идеальных условиях (когда скорость размножения не снижается под действием лимитирующих экологических факторов). Максимальная рождаемость - величина постоянная для данной популяции. В реальных (природных) условиях существования популяции уровень рождаемости определяется различными факторам среды, которые ограничивают скорость появления новых особей. Поэтому для оценки динамики численности популяции используют понятие экологической (реализованной) рождаемости, представляющей увеличение числа особей в популяции в конкретных условиях среды обитания. Экологическая рождаемость - величина непостоянная и сильно варьирует в зависимости от плотности популяции и условий среды обитания.
Различие между максимальной и реализованной рождаемостью можно проиллюстрировать следующим примером. В опытах с мучным хрущаком эти жучки отложили 12 000 яиц (максимальная рождаемость), из которых вылупились только 773 личинки (или 6%)- величина реализованной рождаемости. В общем, для биологических видов, которым не свойственна забота о потомстве (например, многие насекомые, рыбы, земноводные), характерна высокая потенциальная рождаемость и низкая реализованная рождаемость.
Смертность - количество особей в популяции, погибших за определенный период. Понятие смертности противоположно понятию рождаемости. Общее число погибших особей (ΔN) за единицу времени (Δt) называется абсолютной (общей) смертностью. Смертность можно выразить числом особей погибших в единицу времени в расчете на одну особь - удельная смертность (d):
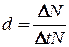
Экологическая (реализованная) смертность - число погибших особей в конкретных природных условиях. Как и экологическая рождаемость, она не постоянна и зависит or особенностей окружающей среды. Теоретическая минимальная смертность - величина постоянная, характеризующая гибель особей (от старости) в идеальных условиях среды (т. е. в отсутствие лимитирующего влияния факторов среды обитания). В конкретных условиях скорость убывания численности популяции определяется гибелью от хищников, полезней и старости.
Часто при описании динамики численности популяции используют понятие выживаемости, т. е. величины, обратной смертности. Если смертность d , то величина выживаемости 1 - d.
Как и рождаемость, смертность и. соответственно, выживаемость у многих организмов в значительной степени варьируют с возрастом. В связи с этим большое значение имеет определение удельной смертности для разных возрастных групп, поскольку это позволяет экологам выяснить механизмы, определяющие общую смертность в популяции. Продолжительность жизни особей популяции можно оценить, используя кривые выживания (рис. 11.1) Откладывая по oси абсцисс возраст особи как процент от общей продолжительности жизни, а по оси ординат - число особей доживших до конкретного возраста, можно сравнить кривые выживания для видов, продолжительность жизни особей которых значительно различается.
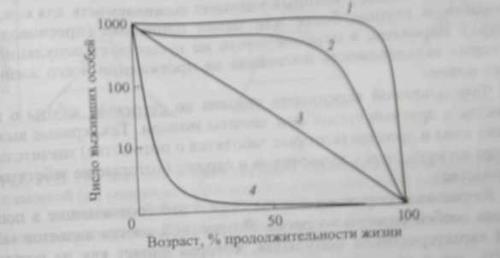
Рис 11.1. Типы кривых выживания; 1 - дрозофила; 2 - человек; 3 - пресноводная гидра; 4 - устрица.
Кривые выживания подразделяются на три общих типа (см. риc. 11.1)
Первый тип (выпуклые кривые 1 и 2) характерен для таких видов в популяции которых наибольшая смертность приходится на конец жизни, т. е. смертность почти до конца жизненного цикла остается низкой и резко повышается только у старых особей. Большинство особей одной популяции имеют примерно одинаковую продолжительность жизни, например, крупные животные.
Другой крайний вариант (сильновогнутая кривая 4) соответствует высокой смертности на ранних стадиях жизненного цикла и повышению выживаемости более взрослых стадий. Этот тип смертности свойственен большинству растений и животных. Максимальная скорость гибели характерна для личиночной фазы развития или в молодом возрасте у животных, а также у многих растений в стадии прорастания семян и всходов. При достижении взрослого состояния организмы становятся более устойчивыми к неблагоприятным воздействиям экологических факторов, и их смертность значительно снижается (и увеличивается выживаемость). Так, при развитии личиночных стадий рыб до половозрелого состояния взрослых особей, доживает, как правило, не более 1...2 % oт общего количества выметанных икринок. У насекомых до половозрелого состояния доживает еще меньше: oт 0,3 до 0,5% от общего количества отложенных яиц.
К промежуточному типу (линия 3) относятся кривые выживания для тех видов, у которых удельная выживаемость для каждой возрастной группы более или менее одинакова (пресноводная гидра). Вероятно, в природе почти не существует популяций, у которых выживаемость постоянна на протяжении всего жизненного цикла.
Форма кривой выживания связана со степенью заботы о потомстве и другими способами защиты молоди. Так, кривые выживания пчел и дроздов (которые заботятся о потомстве) значительно менее вогнуты, чем у кузнечиков и сардин (которые не заботятся о потомстве).
Возрастная структура популяции - это соотношение в популяции особей разного возраста.
Возрастной состав является важной характеристикой популяции, которая влияет как на рождаемость, так и на смертность. Большинство популяций в природе состоит из особей разного возраста и пола.
Упрощенно в популяции можно выделить три экологические возрастные группы:
• предрепродуктивная - молодые особи, еще не достигшие половой зрелости, т. е. не способные участвовать в размножении;
•репродуктивная - половозрелые особи, способные участвовать в размножении;
• пострепродуктивная - старые особи, утратившие способность участвовать в размножении.
Отношение этих возрастов к общей продолжительности жизни в популяции сильно варьирует у разных видов. На количественное соотношение разных возрастных групп в популяции влияют общая продолжительность жизни, время достижения половой зрелости, интенсивность размножения, смертность в разных возрастах. В свою очередь соотношение разных возрастных групп в популяции определяет ее способность к размножению в данный момент и показывает, чего можно ожидать в будущем. Изменение соотношения численности основных возрастных групп в популяциях графически изображается в виде возрастных пирамид (рис. 11.2). В быстрорастущей популяции значительную долю составляют молодые особи (рис. 11.2, а) популяции, численность которой не меняется со временем, возрастной состав более равномерный (рис. 11.2, б), а в популяции, численность которой снижается, будет увеличиваться доля старых особей (рис. 11.2, в).
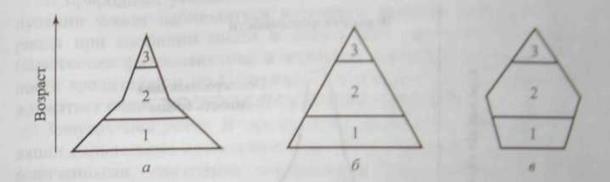
Рис. 11.2. Три типа возрастных пирамид, характеризующие популяции
с высокой (а), умеренной (б) и малой (в) относительной численностью
молодых особей (в % от общей численности популяции):
1 - предрепродуктивная, 2 - репродуктивная, 3- пострепродуктивная возрастная группа
Рост популяции и кривые роста. Если рождаемость в популяции превышает смертность, то наблюдается рост численности популяции.
Каждой популяции и каждому виду в целом свойствен биотический потенциал - максимальная теоретически возможная скорость роста (r) популяции, представляющая собой разность между удельной рождаемостью (b) и удельной смертностью (d):
r = b-d.
Увеличение численности популяции может быть описано кривыми роста двух основных типов - J-образной кривой (экспоненциальный рост) и S-образной кривой (затухающий рост).
Экспоненциальный рост численности популяции характеризуется J-образной кривой роста и происходит когда пищевые пространственные и другие важные жизненные ресурсы  популяции находятся в избытке, а смертность с возрастанием численности особей не увеличивается (рис. 11.3).
популяции находятся в избытке, а смертность с возрастанием численности особей не увеличивается (рис. 11.3).
Уравнение J-образной кривой роста имеет вид
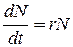
где N - численность популяции; t- время; r - константа скорости роста численности популяции, связанная с максимальной скоростью размножения особи данного вида (биотический потенциал).
Дата добавления: 2016-07-27; просмотров: 18302;
Поиск по сайту
Узнать еще
- I.1.3 ТЕРМОДИНАМИЧЕСКИЙ ПРОЦЕСС. ИЗОПРОЦЕССЫ
- I.2.2 ТЕПЛОЁМКОСТЬ ВЕЩЕСТВА
- II. Поведение веществ в магнитном поле.
- II. Степень насыщенности, определяемая природой связи между атомами углерода.
- II. Установление сегуната Токугава. Социально-экономическое развитие страны: циклы подъема и упадка
- II. Участие Республики Казахстан в международных договорах
- IV. Классификация обязательств РК по международным договорам
- IV. Ликвидация гиповолемии и регуляция водно-солевого обмена

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
