СТРУКТУРА И ФУНКЦИИ ЭКОСИСТЕМ
Общие свойства систем. Центральное понятие в экологии - экосистема отражает основополагающее представление этой науки о том, что природа функционирует как целостная система независимо от того, о какой среде идет речь: пресноводной, морской или наземной. Общая теория сложных систем, к которой относится и изучение интегральных свойств экосистем, начиналась с работ биолога Людвига фон Берталанфи в конце 40-х годов XX в. Системный подход к решению проблем, связанных с окружающей средой, приобретает все большую практическую значимость.
Под системой понимается упорядочение взаимодействующие и взаимозависимые компоненты, образующие единое целое.
Целое - это определенное единство элементов, имеющее свою структуру. Понятие «структура» отражает расположение элементов и характер их взаимодействия.
Системы имеют следующие специфические свойства:
- изоляция;
- интеграция;
- целостность;
- стабильность;
- равновесие;
- управление;
- устойчивость (гомеостаз);
- эмерджентность.
Эмерджентность (от англ. emergence - появление) - универсальная характеристика систем, в том числе экосистем, заключающаяся в том, что свойства системы как целого не являются простой суммой свойств слагающих ее частей или элементов. По мере объединения компонентов в более крупные функциональные единицы, у последних возникают новые свойства, отсутствовавшие на предыдущем уровне (уровне компонентов). Такие качественно новые, эмерджентные, свойства системного уровня организации нельзя предсказать исходя из свойств компонентов составляющих этот уровень или единицу.
Эмерджентные свойства систем возникают в результате взаимодействия компонентов, а не в результате изменения их природы. Учитывая эмерджентные свойства, для изучения целого не обязательно знать все его компоненты, что очень важно для экологии, так как многие экосистемы включают тысячи компонентов-популяций, досконально изучить, которые не представляется возможным. Поэтому на первое место по значимости выступают интегральные свойства целостных сложных экологических систем: суммарная биомасса, продукция и деструкция отдельных трофических уровней, без знания закономерностей, изменения которых нельзя описать поведение всей системы во времени и прогнозировать ее будущее.
Устойчивость саморегулирующихся систем определяет их способность возвращаться в исходное состояние после небольшого отклонения. В этом случае действует принцип Ле Шателье - Бpaунa: при внешнем воздействии, выводящем систему из устойчивого равновесного состояния, равновесие смещается в том направлении, в котором эффект внешнего воздействия ослабляется.
Существование систем немыслимо без прямых и обратных связей. Прямой называют такую связь, при которой один элемент (А) действует на другой (Б) без ответной реакции. Если ответная реакция существует, то говорят об обратной связи (рис. 12.1).
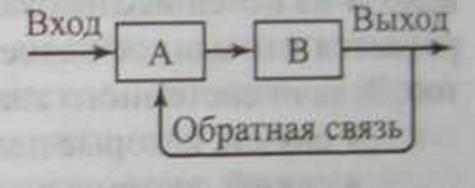
Рис. 12.1 Механизм обратной связи
Этот тип связи играет существенную роль в функционировании экосистем и определяет их устойчивость и развитие. Обратные связи бывают положительные и отрицательные.
Положительная обратная связь обусловливает усиление процесса в одном направлении. Например, после вырубки леса заболачиваются территории, появляются сфагновые мхи (влагонакопители), заболачивание усиливается. Отрицательная обратная связь вызывает в ответ на усиление действия элемента А увеличение противоположной по направлению силы действия элемента Б. Это наиболее распространенный и важный тип связей в природных экосистем. На них прежде всего базируются устойчивость и стабильность экосистем. Пример такой связи - взаимоотношение между хищником и жертвой. Увеличение численности популяции жертв как кормового ресурса создает условия для размножения и увеличения численности популяции хищников. Последние, в свою очередь, начинают более интенсивно уничтожать жертв, уменьшая их численность, и тем самым ухудшают собственные кормовые условия. В менее благоприятных условиях снижается рождаемость в популяции хищника и через некоторое время численность популяции хищников также уменьшается, в результате чего снижается давление на популяцию жертвы. Такая связь позволяет системе сохраняться в состоянии устойчивого динамического равновесия (т. е. саморегулирования).
Обычно различают три вида систем:
1) изолированные - существующие в определенных границах, через которые не происходит обмен веществ и энергии (такие системы создаются только искусственно);
2) закрытые - обменивающиеся со средой только энергией;
3) открытые - обменивающиеся со средой веществом и энергией (это природные экосистемы).
Наиболее важное значение общей теории систем для экологии как науки состоит в том, что она позволила создать новую научную методологию - системный анализ, при которой природные объекты представляются в виде систем. Последние выделяются исходя из целей исследования. С одной стороны, система рассматривается как единое целое, а с другой - как совокупность элементов. Задачи системного анализа состоят в выявлении:
• связей, которые делают систему целостной;
• связей системы с окружающими объектами;
• процессов управления системой;
• вероятности характера поведения исследуемого объекта (прогноз).
Любая система имеет следующие основные параметры:
• границы;
• свойства элементов и системы в целом;
• структуру;
• характер связей и взаимодействия между элементами системы, а также между системой и ее внешней средой.
Границы - наиболее сложная характеристика системы, обусловленная ее целостностью и определяемая тем, что внутренние связи и взаимодействия гораздо сильнее внешних. Последнее обстоятельство определяет устойчивость системы к внешним воздействиям.
Свойства элементов и системы в целом характеризуются качественными и количественными признаками, которые называют показателями.
Структура системы определяется соотношением в пространстве и во времени слагающих ее элементов и их связей. Пространственный аспект структуры характеризует порядок расположения элементов в системе, а временной отражает смену состояний системы во времени (т. е. показывает развитие системы). Структура выражает иерархичность (соподчиненность уровней) и организованность системы.
Характер связей и взаимодействия между элементами системы и системы с внешней средой представляет собой различные формы вещественного, энергетического и информационного обмена. При наличии связей системы с внешней средой границы открыты, в противном случае закрыты.
Экосистема. Живые организмы и их окружение (абиотическая среда обитания) неразделимо связаны друг с другом и находятся и постоянном взаимодействии, образуя экологическую систему (экосистему).
Экосистема - сообщество живых существ и их среда обитания, образующие единое функциональное целое на основе причинно-следственных связей между отдельными экологическими компонентами.
Основные свойства экосистем определяются их способностью осуществлять круговорот веществ и создавать биологическую продукцию, т. е. синтезировать органическое вещество. Природные экосистемы в отличие от искусственных, созданных человеком, при стабильных условиях окружающей среды могут существовать неограниченно долго, так как способны противостоять внешним воздействиям и поддерживать структурно-функциональное постоянство (гомеостаз). Крупные экосистемы включают в себя экосистемы меньшего ранга.
В зависимости от размеров занимаемого пространства экосистемы обычно подразделяют на:
• микроэкосистемы (небольшой водоем, ствол упавшего дерева в стадии разложения, аквариум и т. д.);
• мезоэкосистемы (лес, пруд, озеро, река и т. д.);
• макроэкосистемы (океаны, континенты, природные зоны и т. д.),
• глобальную экосистему (биосфера в целом).
Крупные наземные экосистемы, характерные для определенных географических природных зон, называются биомами (например, тайга, степь, пустыня и т.д.). Каждый биом включат целый ряд меньших по размерам, связанных друг с другом экосистем.
Экосистема состоит из двух основных блоков. Один из них - комплекс взаимосвязанных между собой популяций живых организмов, т. е. биоценоз, а второй - это совокупность факторов среды обитания, т.е. экотоп. Экосистема является функциональной единицей живой природы, включающей биотическую (биоценоз) и абиотическую (среда обитания) части экосистемы, связанные между собой непрерывным круговоротом (обменом) химических веществ, энергию для которых поставляет Солнце (рис. 12.2).
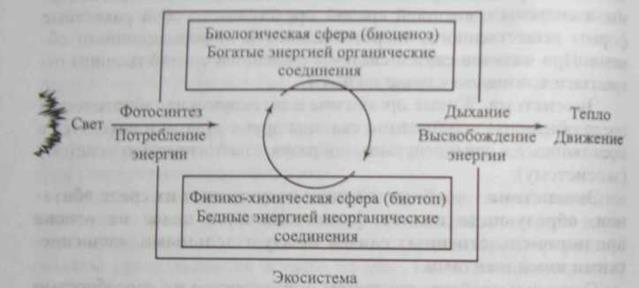
Рис. 12.2. Поток энергии и круговорот химических веществ в экосистеме
Фотосинтезирующие (фотоавтотрофы) организмы (растения, микроводоросли) синтезируют органические вещества из минеральных компонентов почвы, воды и воздуха, используя энергию солнечного света. Образованные в процессе фотосинтеза органические вещества служат растениям источником энергии, необходимым для поддержания своих функций, воспроизводства, а также строительным материалом, из которого они образуют свои ткани (фитомассу). Гетеротрофные организмы (животные, бактерии грибы) в процессе питания используют созданные фотоавтотрофами различные органические соединения для построения своего тела и в качестве источника энергии. В процессе обмена веществ у гетеротрофов происходят высвобождение запасенной химической энергии и минерализация органического вещества до диоксида углерода, воды, нитратов, фосфатов. Поскольку продукты минерализации органического вещества вновь используются автотрофами, возникает постоянный круговорот веществ в экосистеме.
Структура экосистем. Структура любой системы определяется закономерностями в соотношении и связях ее частей. В каждой экосистеме обязательно присутствуют два основных блока элементов: живые организмы и факторы окружающей их неживой среды. Совокупность организмов (растений, животных, микроорганизмов, грибов и т.д.) называют биоценозом или биотой экосистемы. Система взаимоотношений между организмами, а также между биотой и средой обитания, включающей абиотические факторы, определяет структуру экосистемы.
В составе любой экосистемы можно выделить следующие основные компоненты:
- неорганические вещества - минеральные формы углерода, азота, фосфора, вода и другие химические соединения, вступающие в круговорот;
- органические соединения - белки, углеводы, жиры и др.;
- воздушну, водную и субстратную среду, включающую климатический режим (температура и другие физико-химические факторы);
- продуценты - автотрофные организмы, создающие органическую пищу из простых неорганических веществ за счет энергии Солнца (фотоавтрофы), главным образом зеленые растения и одноклеточные микроскопические водоросли в воде, некоторые группы фотосинтезирующих бактерий и хемоавтотрофы, бактерии использующие энергию окислительно-восстановительных реакций (серобактерии, железобактерии и др.);
- консументы - травоядные и хищные гетеротрофные организмы, главным образом животные, которые поедают другие организмы;
- редуценты (деструкторы) - гетеротрофные организмы, преимущественно бактерии и грибы и некоторые беспозвоночные, разлагающие мертвые органические вещества.
Первые три группы компонентов (неорганические вещества, органические вещества, физико-химические факторы) составляют неживую часть экосистемы (биотоп), а остальные - живую часть (биоценоз). Три последних компонента расположенных относительно потока поступающей энергии, представляют собой структуру экосистем (рис. 12.3). Продуценты улавливают солнечную энергию и переводят ее в энергию химических связей органического вещества. Консументы, поедая продуцентов, используют эту энергию для активной жизнедеятельности и построения собственного тела. В результате вся энергия, запасенная продуцентами, оказывается использованной. Редуценты расщепляют сложные органические соединения до минеральных компонентов, пригодных для использования продуцентами (вода, углекислый газ и др.).
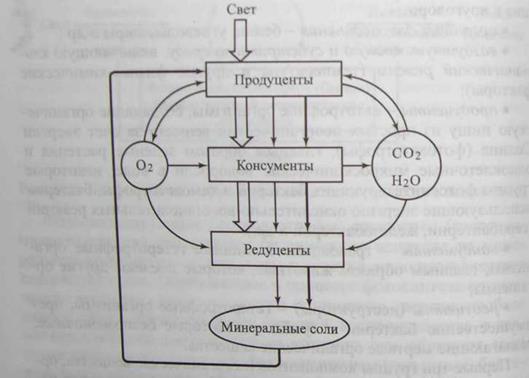
Рис. 12.3. Структура экосистемы, включающая поток энергии (двойная стрелка) и два круговорота веществ: твердых (толстая стрелка) и газообразных (тонкая стрелка)
Таким образом, структуру экосистем образуют три основных группы организмов (продуценты, консументы и редуценты), участвующих в кругoворотax твердых и газообразных веществ, трансформации и использовании энергии Солнца.
Одна из общих черт всех экосистем, будь то наземные, пресноводные, морские или искусственные экосистемы, - это взаимодействие автотрофных (продуценты) и гетеротрофных (консументы и редуценты) организмов, которые частично разделены в пространстве (пространственная структура экосистемы).
Автотрофные процессы (фотосинтез органического вещества растениями) наиболее активно протекают в верхнем ярусе экосистемы, где доступен солнечный свет. Гетеротрофные процессы (биологические процессы, связанные с потреблением органического вещества) наиболее интенсивно протекают в нижнем ярусе, в почвах и осадках, где накапливаются органические вещества.
Система пищевых взаимодействий между организмами формирует трофическую структуру (от греч. trophe - питание), которую для наземных экосистем можно разделить на два яруса:
1) верхний автотрофный ярус (самостоятельно питающийся), или "зеленый пояс", включающий растения или их части, содержащие хлорофилл, в котором преобладают фиксация энергии света, использование проcтыx неорганических соединений и накопление сложных органических соединений, и 2) нижний гетеротрофный ярус (питаемый другими), или «коричневый пояс» почв и осадков, разлагающихся веществ, корней и т. п.. в котором преобладаютиспользование, трансформация и разложение сложных органических соединений.
Функционирование автотрофов и гетеротрофов может быть разделено и но времени, так как использование продукции автотрофных организмов гетеротрофами может происходить не сразу, а с существенной задержкой. Например, в лесной экосистеме фотосинтез протекает преимущественно в кронах деревьев. При этом лишь небольшая часть, продуктов фотосинтеза немедленно и непосредственно перерабатывается гетеротрофами, питающимися листвой и молодой древесиной. Основная масса синтезированного органического вещества (в форме листьев, древесины и запасных питательных веществ в семенах, корнях) в конце концов попадает в почву, где эти вещества относительно медленно используются гетеротрофами. Прежде чем будет использовано все это накопленное органическое вещество, могут пройти многие недели, месяцы, годы или даже тысячелетия (если речь идет oб ископаемых видах топлива).
Следует учитывать, что организмы в природе живут для самих себя, а не для того, чтобы играть какую-либо роль в экосистеме. Свойства экосистем формируются благодаря совокупной деятельности входящих в нее растений и животных. Лишь учитывая это, мы можем понять ее структуру и функции, а также то, что экосистема реагирует на изменения факторов среды как единое целое.
Каждая экосистема характеризуется строго определенной видовой структурой - разнообразием видов (видовым богатством) и соотношением их численности или биомассы. Чем больше разнообразие условий среды обитания, тем больше количество видов в биоценозе. С этой точки зрения самыми богатыми по видовому разнообразию являются, например, экосистемы дождевых тропических лесов и коралловых рифов. Количество видов организмов, населяющих названные экосистемы, исчисляется тысячами. А в экосистемах пустынь существует всего несколько десятков видов.
Видовое разнообразие зависит также от возраста экосистем. В молодых развивающихся экосистемах, возникших, например, на безжизненном субстрате песчаных дюн, горных отвалом, пожарищ, количество видов крайне мало, однако по мере развития экосистем видовое богатство увеличивается.
Из общего числа видов, обитающих в экосистеме, обычно лишь немногие доминируют, т. е. имеют большую биомассу, численность, продуктивность или другие показатели значимости для экосистемы. Большая же часть видов в экосистеме характеризуется относительно низкими показателями значимости.
Не все виды одинаково влияют на свое биотическое окружение. Есть виды-эдификаторы, которые в процессе своей жизнедеятельности формируют окружающую среду для сообщества в целом и без них существование большинства других видов в экосистеме невозможно. Например, ель в еловом лесу является видом-эдификатором, так как создает своеобразный микроклимат, кислую реакцию почвы и специфические условия для развития других видов растений и животных, приспособленных к существованию в данных условиях. При смене елового леса (например, после пожара или вырубки) березовым экотоп на этой территории существенно меняется, что определяет смену всего биологического сообщества экосистемы.
Названия экосистем образуются исходя из важнейших параметров, определяющих характерные условия среды обитания. Так, для наземных экосистем названия включают названия видов-эдификаторов или доминирующих видов растений (ельник-черничник, злаково-разнотравные степные экосистемы и др.).
Функционирование экосистем. Экосистемы являются открытыми системами т. е. такими, которые получают энергию и вещество извне и отдают их во внешнюю среду, поэтому важная составная часть экосистемы - внешняя среда (среда на входе и среда на выходе). Живые организмы, входящие в экосистемы, чтобы существовать, должны постоянно пополнять и расходовать энергию. В отличие от веществ, непрерывно циркулирующих по разным компонентам экосистемы, энергия может быть использована только один раз, т. е. энергия проходит через экосистему в виде линейного потока.
Функциональная схема экосистемы отражает взаимодействие трех основных компонентов, а именно: сообщества, потока энергии и круговорота веществ. Поток энергии направлен только в одну сторону. Часть поступающей солнечной энергии преобразуется биологическим сообществом и переходит на качественно более высокую ступень, трансформируясь в органическое вещество. Но большая часть энергии деградирует: пройдя через систему, выходит в виде низкокачественной тепловой энергии называемой тепловым стоком. Энергия может накапливаться в экосистеме, затем снова высвобождаться или экспортироваться, но она не может использоваться вторично. В отличие от энергии биогенные элементы и вода могут использоваться многократно.
Односторонний поток энергии является результатом действия законов термодинамики. Первый закон термодинамики (закон сохранения энергии) гласит, что энергия может переходить из одной формы (солнечный свет) в другую (потенциальная энергия химических связей в органическом веществе), но она не исчезает и не создается заново, т. е. общее количество энергии в процессах остается постоянным. Второй закон термодинамики (закон энтропии) гласит, что в любых процессах превращения энергии некоторая ее часть всегда рассеивается в виде недоступной для использования тепловой энергии, поэтому эффективность самопроизвольного превращения кинетической энергии (например, света) в потенциальную, (например, в энергию химических связей в органическом веществе) всегда меньше 100 %.
Живые организмы преобразуют энергию, и каждый раз, когда происходит превращение энергии (например, переваривание пищи), часть ее теряется в виде тепла. В конечном счете, вся энергия, поступающая в биотический круговорот экосистемы, рассеивается в виде тепла. Однако живые организмы, населяющие экосистемы, не могут использовать тепловую энергию для совершения работы. Для этой цели они используют энергию солнечной радиации, запасенную в виде химической энергии в органическом веществе, созданном продуцентами в процессе фотосинтеза.
Пища, созданная в результате фотосинтетической активности зеленых растений, содержит потенциальную энергию, которая при использовании ее гетеротрофными организмами превращается в другие формы химической энергии.
Большая часть солнечной энергии, попавшей на землю, превращается в тепловую и лишь очень небольшая её часть (в среднем для земного шара не менее 1%) превращается зелёными растениями в потенциальную энергию химических связей в органическом веществе.
Весь животный мир Земли получает необходимую потенциальную химическую энергию из органических веществ, созданных фотосинтезирующими растениями, и большую её часть в процессе дыхания переводит в тепло, а меньшую вновь преобразует в химическую энергию заново синтезируемой биомассы. На каждом этапе передачи энергии от одного организма к другому её значительная часть рассеивается в виде тепла.
Баланс пищи и энергии для отдельного живого организма можно представить так:
Эп = Эд + Эпр+ Эпв,
где Эп – энергия потребления пищи;
Эд – энергия дыхания;
Эпр – энергия прироста;
Эпв – энергия продуктов выделения.
Выделение энергии в виде тепла в процессе жизнедеятельности у плотоядных животных (хищников) невелико, а у травоядных более значительно. Например, гусеницы некоторых насекомых, питающиеся растениями, выделяют в виде тепла до 70 % поглощенной с пищей энергии. Однако при всем разнообразии величин расходов энергии на жизнедеятельность максимальные траты на дыхание составляют около 90 % всей энергии, потребленной в виде пищи. Поэтому переход энергии с одного трофического уровня на другой в среднем принимаем за 10 % энергии, потребленной с пищей. Эта закономерность известна, как правило, десяти процентов. Из этого правила следует, что цепь питания может иметь ограниченное количество уровней, обычно не более 4-5, пройдя через которые, почти вся энергия оказывается рассеянной.
Пищевые цепи. Внутри экосистемы созданное автотрофными организмами органическое вещество служит пищей (источником энергии и вещества) для гетеротрофов. Типичный пример: животное поедает растение. Это животное, в свою очередь, может быть съедено другим животным, и таким путем может происходить перенос энергии через ряд организмов - каждый последующий питается предыдущим, поставляющим ему сырье и энергию. Такая последовательность организмов называется пищевой цепью, а каждое ее звено - трофическим уровнем. Первый трофический уровень занимают автотрофы (первичные продуценты). Организмы второго трофического уровня называются первичными консументами, третьего - вторичными консументами и т. д.
Главное свойство цепи питания – осуществление биологического круговорота веществ и высвобождение запасенной в органическом веществе энергии.
Представители разных трофических уровней связаны между собой в пищевые цепи процессами односторонне направленной передачи биомассы (в виде пищи, содержащей запас энергии).
Пищевые цепи можно разделить на два основных типа:
1) пастбищные цепи, которые начинаются с зелёного растения и идут дальше к пасущимся животным, а затем к хищникам;
2) детритные цепи, которые начинаются с мелких организмов, питающихся мёртвым органическим веществом, и идут к мелким и крупным хищникам.
Пищевые цепи не изолированы друг от друга, они тесно переплетены в экосистеме образуя пищевые сети.
Экологические пирамиды.Для изучения взаимоотношений между организмами в экосистеме и для графического представления этих взаимоотношений удобнее использовать не схемы пищевых сетей, а экологические пирамиды, основанием которых служит первый трофический уровень (уровень продуцентов), а последующие уровни образуют этажи и вершину пирамиды. Экологические пирамиды можно отнести к трём основным типам:
1) пирамиды численности, отражающие численность организмов на каждом трофическом уровне;
2) пирамиды биомассы, характеризующие общую массу живого вещества на каждом трофическом уровне;
3) пирамиды энергии, показывающие величину потока энергии или продуктивность на последовательных трофических уровнях.
Для графического представления структуры экосистемы в виде пирамиды численности сначала подсчитывают число различных организмов на данной территории, сгруппировав их по трофическим уровням. После таких подсчетов становится очевидно, что численность животных прогрессивно уменьшается при переходе от второго трофического уровня к последующим. Численность растений первого трофического уровня тоже нередко превосходит численность животных, составляющих второй уровень. Два примера пирамид численности показаны на рис. 12.4, где длина прямоугольника пропорциональна количеству организмов на каждом трофическом уровне. Формы пирамид численностей сильно различаются для разных сообществ в зависимости от размеров составляющих их организмов (рис. 12.4).
В пирамидах биомасс учитывается суммарная масса организмов (биомасса) каждого трофического уровня, т. е. показаны количественные соотношения биомасс в сообществе (рис. 12.5). Цифрами обозначено количество биомассы в граммах сухого вещества на 1 м2. В этом случае размер прямоугольников пропорционален массе живого вещества соответствующего трофического уровня, отнесённой к единице площади или объёма. Однако величина биомасс трофического уровня не даёт никакого представления о скорости её образования (продуктивности) и потребления. Например, продуцентам небольших размеров (водоросли) свойственна высокая скорость роста и размножения (увеличение биомассы продуцентов), уравновешенная интенсивным потреблением их в пищу другими организмами (уменьшение биомассы продуцентов). Таким образом, хотя биомасса в конкретный момент может быть малой продуктивность при этом может быть высокой.

Из трех типов экологических пирамид пирамида энергии дает наиболее полное представление о функциональной организации сообщества.

В пирамиде энергии (рис. 12.6), где цифрами обозначено количество энергии (кДж/м2 в год), размер прямоугольников пропорционален энергетическому эквиваленту, т. е. количеству энергии (на единицу площади или объема), прошедшей через определенный трофический уровень за конкретный период. Пирамида энергии отражает динамику прохождения массы пищи через пищевую (трофическую) цепь, что принципиально отличает её от пирамид численности и биомассы отражающих статическое состояние экосистемы (количество организмов в данный момент).
Продуктивность экосистем –образование органического вещества в виде биомассы животных, растений и микроорганизмов, составляющих биотическую часть экосистемы, в единицу времени на единицу площади или объема. Способность создавать органическое вещество (биологическая продуктивность) - одно из важнейших свойств организмов, их популяций и экосистем в целом.
За счет энергии света при фотосинтезе создается основная, или первичная, продукция экосистемы. Первичная продуктивность – это скорость, с которой солнечная энергия усваивается продуцентами (растениями) в процессе фотосинтеза, накапливаясь в форме органических веществ.Иными словами, это величина скорости прироста биомассы растений.
Принято выделять четыре последовательные стадии процесса производства органического вещества:
1) валовая первичная продуктивность - общая скорость фотосинтеза, т. е. скорость образования всей массы органических веществ продуцентами, включая и то количество органического вещества, которое было израсходовано продуцентами на поддержание деятельности (РG);
2) чистая первичная продуктивность - скорость накопления органического вещества в растительных тканях за вычетом того органического вещества, которое было синтезировано растениями и использовано на поддержание своей жизнедеятельности (РN);
3) чистая продуктивность сообщества - скорость накопления органического вещества, не потребленного гетеротрофами (животными и бактериями), в сообществе за конкретный период (например, прирост биомассы растений к концу летнего сезона).
4) вторичная продуктивность - скорость накопления энергии (в виде биомассы) на уровне консументов (животных), которые не создают органическое вещество из неорганических (как в случае фотосинтеза), а лишь используют органические вещества, полученные с пищей, часть из них расходуя на поддержание жизнедеятельности а остальные превращая в собственные ткани.
Высокие скорости продукции органического вещества встречаются при благоприятных факторах окружающей среды, особенно при поступлении дополнительной энергии извне, уменьшающей собственные затраты организмов на поддержание жизнедеятельности. Например, в прибрежной зоне моря дополнительная энергия может поступать в форме энергии приливов, приносящих малоподвижным организмам частицы органического вещества.
Дня наглядного представления региональных особенностей функционирования биосферы на рис. 12.7 приведена модель продуктивности крупных экосистем биосферы в виде турбины, работающей от потока солнечных лучей. Ширина колеса турбины для суши соответствует проценту суши в конкретной природной зоне, ширина колеса для моря взята произвольно. Лопатки этой модельной турбины (виды растений в конкретной экосистеме) воспринимают солнечный свет в процессе фотосинтеза и обеспечивают энергией все жизненные процессы в экосистемах. При этом сухопутная турбина имеет наибольшее количество лопаток (видов растений) в области тропиков, где 40 тыс. видами растений может вырабатываться годичная биологическая продукция в 1011 т органического вещества. В тропических экосистемах суши в среднем за год вновь создается около 800 г/м2 углерода. Морские экосистемы (рис. 12.7) наиболее продуктивны в умеренных бореальных областях, где в год образуется около 200 г углерода на 1 м2.
Величина биологической продуктивности является определяющей для большинства систем классификации водоемов по уровню трофности, т. е. обеспеченности питательными веществами для развития биоценоза. Уровень трофности водоема определяется по содержанию основного фотосинтетического пигмента (хлорофилла), по величине общей биомассы и по скорости продукции органического вещества. Согласно этой классификации выделяют четыре типа озер: олиготрофные, эвтрофные, мезотрофные и гипертрофные (табл. 12.1).

В предложенной системе классификации уровень биологической продуктивности (трофность) водоёмов тесно связан с абиотическими факторами (глубина, цветность, прозрачность водоема, наличие кислорода в придонных слоях воды, кислотность воды (рН), концентрация биогенных элементов и пр.), с географическим положением водоема и характером водосборного бассейна.

Олиготрофные водоёмы (от греч.- незначительный, бедный) содержат незначительное количество биогенных веществ, имеют высокую прозрачность низкую цветность, большую глубину. Фитопланктон в них развит незначительно, так как автотрофные организмы не обеспечены минеральным питанием, главным образом азотом и фосфором. Синтезированное в водоёме органическое вещество (автохтонное вещество) практически полностью (до90..95%) подвергается биохимическому распаду. В результате в донных отложениях количество органического вещества небольшое, поэтому в придонных слоях воды содержание кислорода высокое. В водоеме преобладают пастбищные трофические цепи, микроорганизмов мало и деструкционные процессы выражены слабо. Подобные озера характеризуются большими размерами и большой глубиной.
Эвтрофные водоемы (от греч. eutrophia хорошее питание) характеризуются повышенным содержанием биогенных элементов (азот и фосфор), поэтому фитопланктон обеспечен минеральным питанием и интенсивность продукционных процессов высокая. С увеличением степени эвтрофирования уменьшаются прозрачность и глубина зоны фотосинтеза. В верхних слоях воды часто возникает избыток кислорода благодаря высокой скорости фотосинтеза, тогда как в придонных слоях воды - значительный дефицит кислорода из-за использования его микроорганизмами в процессах окисления органического вещества. В водоеме все большее значение приобретают детритные цепи питания.
Мезотрофный тип (от греч. mesos - средний) - промежуточный тип водоемов между олиготрофным и эвтрофным. Обычно мезотрофные водоемы возникают из олиготрофных и превращаются в эвтрофные. Во многих случаях этот процесс связан с эвтрофикацией - повышением уровня первичной продукции вод благодаря увеличению в них концентрации биогенных элементов, главным образом азота и фосфора. Поступление биогенных элементов в водоемы увеличивается в результате смывания с полей удобрений, а также попадания в них промышленных и коммунальных стоков.
Гипертрофные водоемы (от греч. hyper - над, сверх) характеризуются очень высоким уровнем первичной продукции и, как следствие высокой биомассой фитопланктона. Прозрачность и содержание кислорода в водоемах минимальные. Содержание большого количества органического вещества приводит к массовому развитию микроорганизмов, которые преобладают в биоценозе.
Гомеостаз экосистемы.Экосистемы, подобно входящим в их состав популяциям и организмам, способны к самоподдержанию и саморегулированию. Гомеостаз (от греч. подобный, одинаковый) – способность биологических систем противостоять изменениям и сохранять динамическое относительное постоянство состава и свойств. Нестабильность среды обитания в экосистемах компенсируется биоценотическими адаптивными механизмами.
Наряду с потоками энергии и круговоротами веществ экосистему характеризуют развитые информационные сети, включающие потоки физических и химических сигналов, связывающих все части системы и управляющих ею как единым целым. Поэтому можно считать, что экосистемы имеют и кибернетическую природу.
В основе гомеостаза лежит принцип обратной связи, который можно продемонстрировать на примере зависимости плотности популяции от пищевых ресурсов. Обратная связь возникает, если «продукт» (численность организмов) оказывает регулирующее влияние на «датчик» (пищу). В данном примере количество пищевых ресурсов определяет скорость прироста популяции. При отклонении плотности популяции от оптимума в ту или иную сторону увеличивается рождаемость или смертность, в результате чего плотность приводится к оптимуму. Такая обратная связь, уменьшающая отклонение от нормы, называется отрицательной обратной связью.
Помимо систем обратной связи стабильность экосистемы обеспечивается избыточностью функциональных компонентов. Например, если в сообществе имеется несколько видов автотрофов, каждый из которых характеризуется своим температурным оптимумом, то при колебаниях температуры окружающей среды скорость фотосинтеза сообщества в целом будет оставаться неизменной.
Гомеостатические механизмы действуют в определенных пределах, за которыми уже ничем не ограничиваемые положительные обратные связи приводят к гибели системы, если невозможна дополнительная настройка. По мере нарастания стресса система, продолжая оставаться управляемой, может оказаться неспособной к возвращению на прежний уровень.
Область действия отрицательной обратной связи можно изобразить в виде гомеостатического плато (рис. 12.8). Оно состоит из ступенек; в пределах каждой ступеньки действует отрицательная обратная связь. Переход со ступеньки на ступеньку может произойти в результате изменения в «датчике». Так, увеличение или уменьшен<
Дата добавления: 2016-07-27; просмотров: 20969;
Поиск по сайту
Узнать еще
- a-спираль b-складчатая структура
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I.2. Антигены системы АВ0. Генетика. Структура
- II. Митохондрии (строение и функции)
- II. Признаки, ресурсы и функции власти.
- II.1.2. Гены. Структура антигенов
- III. Функции отдела по делам ГОЧС и ВМР
- III. Функции политологии. Возрастание роли политических знаний в жизни общества.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
