РАЗВИТИЕ И ЭВОЛЮЦИЯ ЭКОСИСТЕМ
Отдельные виды организмов не только сами приспосабливаются к экологической среде, но и в ходе своей совместной деятельности в экосистемах приспосабливают геохимическую среду экосистемы к своим потребностям. Таким образом, сообщества организмов и среда их обитания развиваются как единое целое.
Развитие экосистемы, или экологическая сукцессия (от лат. successio - преемственность, наследование), - это изменение во времени видовой структуры и биоценотических процессов. В ходе сукцессии изменяется окружающая среда под действием сообщества живых организмов и в экосистемах одни виды заменяются другими. В отсутствие внешних нарушающих процессов сукцессия является направленным, закономерным и предсказуемым процессом изменения экосистемы в сторону большей стабильности и сбалансированности. Сукцессия - результат изменения развивающимся сообществом организмов физико-химической среды и взаимодействий типа «конкуренция-сосуществование» на популяционном уровне. Таким образом, сукцессия контролируется сообществом, несмотря на то, что физическая среда определяет характер и скорость изменений, а часто и ограничивает пределы развития. Если сукцессионные изменения определяются преимущественно внутренними взаимодействиями, то говорят об аутогенной (т. е. саморазвивающейся) сукцессии. Если же на процессы сукцессионных изменений регулярно воздействуют или контролируют их факторы окружающей среды (штормы, пожары и т. д.), то такую сукцессию называют аллогеннои (порожденной извне).
Процесс сукцессии начинается в результате изменений фундаментальных свойств среды обитания, происходящих под влиянием комплекса факторов. Такие факторы могут быть естественными (отступление ледников, наводнения, землетрясения, извержения вулканов, пожары) или антропогенными (расчистка лесных угодий, распашка участков степи, создание водохранилищ, загрязнение среды).
Обычно различают первичные и вторичные, автотрофные т гетеротрофные, аллохтонные и автохтонные сукцессии.
Первичные сукцессии начинаются всякий раз, когда заселяются пустынные места обитания. Это могут быть голые скалы, остывшая вулканическая лава, песчаные дюны. Первыми, как правило, на таких пустынных местах появляются лишайники, за ними следуют мхи и неприхотливые однолетние травы, затем двухлетние и многолетние растения, кустарники и, наконец, - деревья. Течение этого процесса определяется скоростью накопления почвенного слоя и повышения влажности.
Таким образом, первичная сукцессия на суше начинается с почвообразования, проявляющегося в физическом и химическом выветривании, ведущем к разрыхлению поверхности материнских горных пород, таких как базальты, гнейсы, граниты, известняки, песчаники, сланцы. Этот слой, образовавшийся в результате выветривания, постепенно заселяется автотрофными микроорганизмами и лишайниками, которые преобразуют субстрат и обогащают его органическими веществами. В результате жизнедеятельности лишайников в первичной почве накапливаются важнейшие элементы питания растений: фосфор, кальций, калий и др. На этой почве теперь могут поселиться травянистые растения, способные сформировать растительные сообщества, определяющие основу биогеоценоза.
Примером первичной сукцессии могут служить исторически зафиксированные процессы восстановления экосистемы на острове Кракатау в Индонезийском море. В ходе катастрофического извержения вулкана Кракатау в конце XIX в. большая часть острова была уничтожена взрывом, а сохранившаяся часть была покрыта 60-метровым слоем лавы и горячего вулканического пепла. Все живое на острове было уничтожено. Год спустя прибывшие на остров ученые обнаружили несколько видов травянистой растительности и один вид пауков. Через 25 лет на острове обитало уже 202 вида животных и растений; через 36 лет - 621 вид. А через 50 лет там насчитывалось 880 видов организмов, к тому же на острове рос молодой тропический лес.
Вторичная сукцессия. Характерна, для мест, где ранее существовали биологические сообщества, которые были частично или полностью разрушены в результате естественных (пожары, наводнения) либо антропогенных причин (вырубки, загрязнение среды), но в почве или донных отложениях сохранились семена растений, споры бактерий и грибов.
Примером вторичных сукцессионных изменений может служить зарастание заброшенных сельскохозяйственных угодий или восстановление лесных массивов (рис. 13.1). В лесной зоне на и исходно безжизненной территории сначала появляются организмы - пионеры, например мхи или корковые водоросли, лишайники. За ними следует растительность, представленная в основном травами, а затем появляются кустарники и деревья-пионеры (береза, осина ива). Для последних характерен быстрый рост. Под пологом этих деревьев создаются условия для поселения теневыносливой ели, которая постепенно догоняет в росте стареющие лиственные деревья и выходит в верхний ярус, вытесняя светолюбивые виды деревьев и кустарников. Соответственно на каждой стадии сукцессии меняется и животный мир.
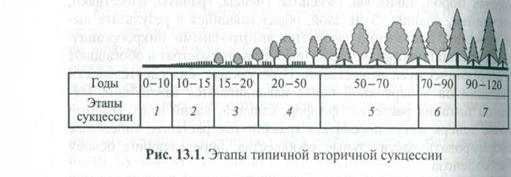
Смена стадий сукцессии, которые называются сериями, идет в соответствии с определенными правилами. Так, каждая предыдущая стадия сукцессии подготавливает среду для возникновения последующей. При этих изменениях в экосистеме постепенно возрастают видовое многообразие и ярусность. Вслед за растениями в сукцессию вовлекаются представители животного мира, а развивающийся биоценоз становится более богатым видами. Цепи питания в нем усложняются, развиваются и превращаются в сети питания.
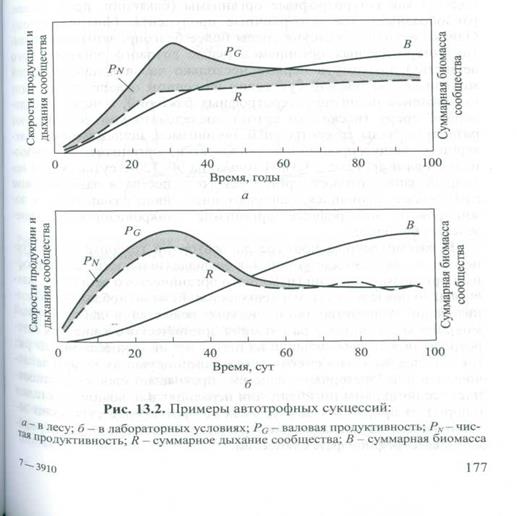
Различают также автотрофные и гетеротрофные сукцессии. Автотрофная сукцессия характерна для случаев заселения новых территорий или создания водохранилищ. Она начинается с несбалансированного сообщества, в котором величина суммарной валовой первичной продуктивности (Р) превышает величину суммарного дыхания сообщества (R), т. е. на начальной cтадии автотрофной сукцессии скорость образования органического вещества в ходе фотосинтеза существенно превышает скорость его расходования в процессе дыхания всех обитающих в экосистеме организмов (Р / R > 1). ). По мере развития экосистемы скорости их процессов сравниваются (рис. 13.2) и отношение P/R стремится к 1, что свидетельствует о сбалансированности процессов синтеза и распада органического вещества в зрелой стадии развития экосистемы. Как видно из рис. 13.2, общие закономерности автотрофной сукцессии можно наблюдать и в природной лесной экосистеме, и в условиях лабораторных микроэкосистем (микрокосмов): величины чистой первичной продуктивности органического вещества возрастают на ранних стадиях развития сообщества, но убывают - на поздних. Однако скорость изменений этих важнейших интегральных характеристик состояния экосистемы (продукция, деструкция) и время, необходимое для достижения стабильного состояния экосистем, варьируют.
Гетеротрофная сукцессия начинается тогда, когда существует запас доступного гетеротрофным организмам мёртвого органического вещества, и затраты на дыхание сообщества (R) на начальной стадии развития сообщества превалируют над валовой первичной продуктивностью (P/R < 1). Этот тип сукцессии наблюдается в условиях, когда в начале сукцессии продуценты практически отсутствуют, а доминирующими в биоценозе являются гетеротрофы.
Примером подобной сукцессии может служить развитие сообщества водных организмов в лабораторных условиях на сенном отваре (рис. 13.3). В начальный момент лабораторной модели сукцессии в стерильный сенной отвар вносят одну каплю воды из водоема (пруда, реки, озера). В этой исходной капле воды находятся как гетеротрофные организмы (бактерии, простейшие, ракообразные), так и первичные продуценты (фитопланктон). Однако исходные условия среды более благоприятны для развития гетеротрофных организмов (запас готового органического вещества), поэтому в первые несколько часов в лабораторной модели гетеротрофной сукцессии на сенном отваре происходит интенсивное развитие гетеротрофных бактерий, а через определенное время (несколько суток) последовательно доминируют различные виды гетеротрофных организмов, использующих бактерии в пищу: простейшие, коловратки, организмы микрозоопланктона и др. (рис. 13.3). И только на 90-120-е сутки, когда исходный запас готового органического вещества в лабораторном микрокосме закончился, начинают интенсивно развиваться и доминировать автотрофные организмы (микроводоросли, сине-зеленые бактерии).
В рассмотренном примере лабораторной гетеротрофной сукцессии количество доступной для жизнедеятельности гетеротрофных организмов энергии (исходного органического вещества) максимально вначале, а затем уменьшается. Если не добавлять дополнительное количество органического вещества в лабораторный микрокосм, то по мере разрушения органического вещества гетеротрофами и высвобождения из него энергии сукцессионный ряд (ряд последовательно сменяющихся доминирующих видов) заканчивается и лабораторная экосистема прекращает своё существование: все организмы погибают или переходят в покоящиеся стадии (споры). В природных экосистемах гетеротрофные сукцессии заканчиваются не распадом биологического сообщества, а переходом к автотрофной фазе сукцессии.
Другим примером гетеротрофной сукцессии может быть река, загрязненная большим количеством органических отбросов (стоки с ферм). Избыточное органическое вещество, поступающее в экосистему, активно используется гетеротрофами. При этом в результате жизнедеятельности автотрофных организмов создается значительно меньше органического вещества, чем его разрушается в экосистеме.

Процесс сукцессионных изменений в экосистеме всегда направлен в сторону выравнивания или сбалансированности процессов новообразования и разрушения органического вещества. Если процесс сукцессионных изменений не нарушается вмешательством извне (вырубки, пожары, выпас скота, антропогенное воздействие), то в конце концов образуется устойчивое климаксное сообщество. Дальнейшие изменения в таком сообществе происходят, и оно может существовать неопределенно долгое время Климакс экосистем определяется местными климатическими условиями. При отсутствии лимитирующих факторов все
сукцессионные изменения заканчиваются климаксом. Тайга, тундра, саванна, степь. Пустыня – примеры климаксных экосистем, характерных для конкретных природных зон. Однако реальная возможность произрастания тех или иных видов в данном климате зависит от почвенных условий. Поэтому, хотя все сукцессии в данной климатической области стремятся к одному и тому же климаксу, тип почвы, рельеф местности, расположение водоемов могут помешать сукцессии дойти до конца. Подобного рода климакс, называется почвенным или эдафическим.
По мнению некоторых исследователе, все сукцессии в данном климатогеографическом районе при наличии достаточного времени приведут к одному и тому же климаксу (моноклимаксная теория). По мнению других, такая возможность существует лишь теоретически, а в реальных условиях сукцессия прекращается, достигая различных эдафических климаксов (поликлимаксная теория).
Например, типичная климаксная экосистема Подмосковья - дубрава с подлеском орешника и с доминированием осоки в травяном покрове. Однако практически на всей территории Московской области наземные лесные сообщества испытывают частые антропогенные нарушения, и в результате климаксная экосистема (дубрава) занимает относительно небольшую часть территории области.
Для завершения большинства сукцессии требуются сотни и тысячи лет. Например, в лесной зоне умеренного пояса Европейской части России для завершения сукцессии, начавшихся на голом месте, требуется от 200 до 1000 лет. Длительность конкретной сукцессии зависит от местных условий. В водоемах с сильным течением или на крутых склонах гор в результате отмирания животных и растений откладывается относительно меньше органического вещества, чем на более ровных участках поверхности земли и в водоемах с медленным течением. В итоге скорость сукцессии резко замедляется.
В других случаях на сообщество периодически действуют внешние силы, нарушающие течение сукцессии (пожары, вырубки, наводнения и т. д.). Каждое такое нарушение отбрасывает сообщество в своем развитии на предыдущие стадии сукцессии. Если при этом растительность и почва не полностью уничтожены, то начинаются вторичные сукцессии. В итоге климаксное сообщество будет занимать лишь небольшие участки территории, где никаких катаклизмов и антропогенных вмешательств не происходило.
Изменение сообщества в ходе сукцессии. По мере замены одного биологического сообщества другим в ходе сукцессии, прежде всего изменяется его видовой состав. Среди видов различают виды доминанты (господствующие в сообществе) и виды-эксплеренты, которые конкурентно слабее доминантов и тем не менее быстро захватывают участки экосистемы с нарушенным растительным покровом.
Среди номинантов выделяют виды-эдификаторы, в наибольшей степени формирующие характерную для сообщества среду. Существуют еще виды-ценофобы (виды-сорняки), которые поселяются на участках, лишенных растительности, еще до появления пионерных видов сукцессионного ряда. Ценофобы поселяются на пустырях, свалках и других местах, где растительность уничтожена человеком. От пионерных видов ценофобы отличаются тем, что их присутствие не обязательно для последующих стадий сукцессии. В любом сообществе время от времени появляются участки, лишенные растительного покрова, поэтому растения ценофобы могут присутствовать на всех стадиях сукцессии.
В ходе сукцессии более простые биологические сообщества, характерные для начальных стадий развития сукцессии (с низким видовым разнообразием, простой пространственной структурой, несбалансированным приходом и расходом энергии и вещества), сменяются сообществами с более сложной структурой.
Обычно в ходе сукцессии увеличивается биомасса сообщества, сложным образом изменяется видовое разнообразие (на ранних стадиях оно быстро растет, на промежуточных достигает одного или нескольких максимумов, а по достижении климакса снижается). Виды, являющиеся r-стратегами, первыми захватывают освободившиеся участки территорий с избытком пищевых ресурсов, но позднее, когда свободных ресурсов становится меньше и усиливаются конкурентные взаимоотношения между видами, r-стратеги вытесняются К-стратегами.
Рассмотрим важнейшие закономерности изменения во времени, т. е. динамики, основных интегральных структурно-функциональных характеристик состояния экосистемы (валовой и чистой первичной продуктивности PG и PN) деструкции (R), биомассы (В), численности видов (N) в ходе автотрофной аутогенной (саморазвивающейся) сукцессии. Причем изменения состояния экосистемы в ходе автотрофной сукцессии представим в отношении энергетического баланса, круговоротов веществ, видовой структуру сообщества и стабильности экосистемы в целом.
Изменения в энергетическом балансе:
• возрастают биомасса (В) и количество мертвого органически го вещества (детрита);
• возрастает валовая первичная продуктивность (PG) - скорость образования органического вещества в ходе фотосинтеза;
• увеличивается дыхание сообщества (R) - скорость разрушения органического вещества в ходе процессов жизнеобеспечения организмов;
• уменьшается величина чистой продуктивности (PN) органического вещества (PN = PG - R);
• увеличивается сбалансированность процессов синтеза и распада органического вещества - величина коэффициента (PN/R) стремится к 1.
Изменения круговоротов биогенных элементов:
• становятся все более замкнутыми в пределах экосистемы (т. е. повышается эффективность трансформации и повторного использования веществ в сообществе);
• увеличиваются время оборота и запас биогенных элементов в экосистеме (скорость оборота элементов снижается, так как большая их часть заключена в химически связанном виде в составе биомассы или детрита).
Изменения видовой структуры сообщества:
• становится другим видовой состав сообщества (пионерные виды, первыми заселяющие безжизненные территории, подготавливают условия для существования последующих группировок видов);
• возрастает видовое разнообразие;
• на ранних стадиях развития экосистемы доминируют r-стратеги (с J-образным характером роста популяции), а на более поздних стадиях - К-стратеги (с S- образным характером роста популяции);
• кроме того, усложняются и удлиняются жизненные циклы организмов, составляющих биоценоз экосистемы.
Изменение стабильности экосистемы выражается в возрастании устойчивости биологического сообщества к различным воздействиям извне в результате адаптивных структурных изменений (изменения видовой структуры направлены на уменьшение негативного воздействия).
Существует классификация различных типов экосистем, основанная на интенсивности метаболизма сообществ (совокупности процессов использования вещества и энергии в ходе активной жизнедеятельности биосистем) и на сопоставлении сбалансированности интегральных процессов новообразования и разрушения органического вещества.
На ранних стадиях сукцессии скорость валовой первичной продуктивности (Р) и общего дыхания сообщества (R) существенно различаются: P/R > 1 при автотрофной сукцессии и P/R<1 при гетеротрофной сукцессии. В обоих случаях величина P/R приближается к единице по мере развития сукцессии. Иными словами, в зрелой, климаксной, экосистеме наблюдается динамическое равновесие между связанной энергией (продуктивность органического вещества - Р) и энергией, затрачиваемой на поддержание жизнедеятельности организмов (т. е. суммарным дыханием биологического сообщества). Таким образом, отношение P/R является функциональным показателем стадии сукцессии или относительной зрелости экосистемы.
Графически равновесные состояния экосистем при разных уровнях продуктивности и дыхания в сообществах можно изобразить в виде прямой, составляющей угол 45° с осями координат Р и R (рис. 13.4). На диаграмме показано положение разных экосистем в системе классификации, основанной на метаболизме сообществ.
Выше диагональной линии на рис. 13.4 располагается область существования автотрофных сообществ, так как валовая первичная продуктивность (Р) превышает дыхание (R) и P/R больше единицы.
Справа от диагонали видим обратную ситуацию: P/R меньше единицы, и там располагаются гетеротрофные сообщества. В последнем случае биотические сообщества существуют за счет органического вещества, получаемого извне, или за счет его предварительного накопления. Стрелками показано направление сукцессионных изменений в экосистемах в ходе автотрофной и гетеротрофной сукцессии. Климаксные сообщества, расположенные вдоль диагонали, в среднем за год потребляют примерно столько же энергии, сколько создают ее за это же время, поэтому их можно считать мстаболическими климаксами.
Климаксные сообщества обладают большим видовым разнообразием, сложной трофической структурой, сбалансированностью процессов продукции и распада органического вещества, поэтому они способны в гораздо большей степени противостоять негативным воздействиям физических факторов (температур влажность) и даже некоторым видам химических загрязнений по сравнению с сообществами на ранних сукцессионных стадиях. Однако молодое биотическое сообщество способно продуцировать новую биомассу с гораздо большей скоростью, чем сообщество, находящееся в стадии климакса. Поэтому с экономической точки зрения наиболее эффективно использование возобновимых природных ресурсов (биомассы) в молодых развивающихся экосистемах, находящихся на ранних стадиях сукцессии.
Устойчивость биотического сообщества, находящегося на стадии климакса, его способность противостоять воздействию физических факторов являются очень важными и весьма желательными для человека свойствами. Во имя нашего собственного благополучия большинство природных ландшафтов должно быть сохранено вместе с формировавшимися на протяжении веков биологическими сообществами.

Контрольные вопросы и задания
1. Дайте определение экологической сукцессии. Какие бывают виды сукцессии?
2. Приведите примеры первичных и вторичных, автотрофных и гетеротрофных сукцессии.
3. Какие изменения происходят в экосистемах в ходе сукцессионного развития?
4. Охарактеризуйте климаксную экосистему.
Дата добавления: 2016-07-27; просмотров: 9934;
Поиск по сайту
Узнать еще
- Arthropoda.Систематика.Блохи.Виды блох.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское и эпидемиологическое значение.Меры борьбы.
- Arthropoda.Систематика.Мошки,мокрецы,слепни,оводы.Географическое распространение.Морфология,развитие,патогенное действие.Медицинское значение,меры борьбы.
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- Cоциально-политическое развитие Киевской Руси. Деятельность первых древнерусских князей.
- II. Политическое развитие страны: реформы и их последствия
- II. Установление сегуната Токугава. Социально-экономическое развитие страны: циклы подъема и упадка
- IV этап: 1980 г. – по настоящее время. Интенсивное развитие стратегического маркетинга
- Автохтонное развитие

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
