БІОФІЗИЧНІ ОСНОВИ ЗОРОВОЇ РЕЦЕПЦІЇ
Око, як відомо, становить складну оптичну систему, яка до того ж є джерелом інформації і про деякі незорові захворювання.
Розглянемо будову ока (рис. 6.42). Очне яблучко мас майже сферичну форму діаметром 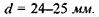 Зовнішня білкова оболонка - склера 1-у передній частині перетворюється в прозору, опуклу роговицю 2. Роговиця є частиною ока, що має найбільшу заломлюючу силу (показник заломлення
Зовнішня білкова оболонка - склера 1-у передній частині перетворюється в прозору, опуклу роговицю 2. Роговиця є частиною ока, що має найбільшу заломлюючу силу (показник заломлення 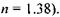 До склери прилягає судинна оболонка З, яка на передній частині пігментована і утворює практично непрозору райдужну оболонку 4. Пігментні клітини визначають колір райдужної оболонки, тобто "колір ока". Райдужна оболонка має отвір - зіницю, що відіграє роль апертурної діафрагми. Діаметр зіниці за допомогою м'язових волокон змінюється від
До склери прилягає судинна оболонка З, яка на передній частині пігментована і утворює практично непрозору райдужну оболонку 4. Пігментні клітини визначають колір райдужної оболонки, тобто "колір ока". Райдужна оболонка має отвір - зіницю, що відіграє роль апертурної діафрагми. Діаметр зіниці за допомогою м'язових волокон змінюється від 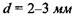 до
до 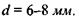
До зіниці прилягає кришталик 5 - прозоре тіло, подібне до двоопуклої лінзи. Показник заломлення змінний і становить 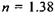 на периферії і
на периферії і 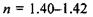 в центрі. Між роговицею та кришталиком розташована передня камера 6, заповнена водянистою вологою. Внутрішню частину ока заповнює прозора студениста маса - скловидне тіло 7, що має такий самий показник заломлення, як і водяниста волога, а саме:
в центрі. Між роговицею та кришталиком розташована передня камера 6, заповнена водянистою вологою. Внутрішню частину ока заповнює прозора студениста маса - скловидне тіло 7, що має такий самий показник заломлення, як і водяниста волога, а саме: 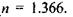 До судинної оболонки в задній частині, що має назву окового дна, прилягає - сітківка, або ретина 8, яка відіграє роль світлосприймаючого екрану. Заломлюючі середовища ока - роговиця, волога передньої камери, кришталик, скловидне тіло - утворюють разом центровану оптичну систему (ЦОС) з головною віссю (ОО), що проходить через центри роговиці, зіниці та кришталика.
До судинної оболонки в задній частині, що має назву окового дна, прилягає - сітківка, або ретина 8, яка відіграє роль світлосприймаючого екрану. Заломлюючі середовища ока - роговиця, волога передньої камери, кришталик, скловидне тіло - утворюють разом центровану оптичну систему (ЦОС) з головною віссю (ОО), що проходить через центри роговиці, зіниці та кришталика.
Око має також зорову вісь 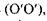 яка проходить через центри кришталика та жовтого п'ятна - найчутливішої частини сітківки.
яка проходить через центри кришталика та жовтого п'ятна - найчутливішої частини сітківки.
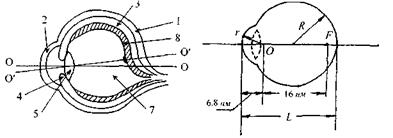
Рис. 8.42. Будова ока. Рис. 8.43. Приведене око в мо-
делі Вербицького.
Для ЦОС ока оптична сила становить 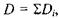 де
де  оптичні сили елементів цієї системи. Оптичні сили світло-сприймаючих
оптичні сили елементів цієї системи. Оптичні сили світло-сприймаючих  елементів ока становлять
елементів ока становлять 
Таким чином, 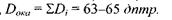
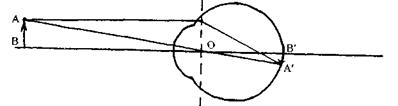
Рис. 6.44. Побудова зображення в приведеному оці.
Для побудови зображення на сітківці ока використовують так зване приведене око.Розглянемо приведене око у моделі Вербицького (рис. 6.43). Радіус сфери 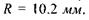 Радіус передньої заломлюючої поверхні
Радіус передньої заломлюючої поверхні 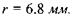 Довжина лінзи вздовж вісі
Довжина лінзи вздовж вісі 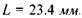 Головний фокус
Головний фокус 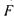 знаходиться на відстані
знаходиться на відстані 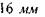 від оптичного центра. Показник заломлення лінзи
від оптичного центра. Показник заломлення лінзи 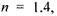 а оптична сила
а оптична сила 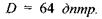
Побудова зображення у приведеному оці здійснюється за тими правилами, як і для однієї лінзи: предмет знаходиться за подвійним фокусом, зображення - на задній поверхні приведеного ока - дійсне, зменшене (рис. 6.44). Віддалені предмети, що знаходяться на різній відстані від ока, повинні давати на сітківці однаково різкі зображення. Тому око є самонастроювальною системою.
Відповідно до формули лінзи 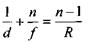 кожному значенню
кожному значенню 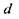 повинно відповідати певне значення
повинно відповідати певне значення 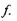 Але
Але 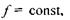 тому зменшення
тому зменшення 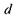 повинно компенсуватися зміною радіуса кривизни
повинно компенсуватися зміною радіуса кривизни 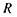 кришталика або фокусної відстані
кришталика або фокусної відстані 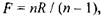 тобто із зменшенням
тобто із зменшенням 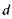 збільшується кривизна кришталика або, що теж саме, зменшується радіус кривизни
збільшується кривизна кришталика або, що теж саме, зменшується радіус кривизни 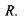 Таким чином, око здатне до акомодації. Акомодація - це здатність ока утворювати на сітківці однаково різкі зображення по-різному віддалених предметів. Акомодація здійснюється за допомогою зміни оптичної сили
Таким чином, око здатне до акомодації. Акомодація - це здатність ока утворювати на сітківці однаково різкі зображення по-різному віддалених предметів. Акомодація здійснюється за допомогою зміни оптичної сили 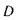 ока за рахунок зміни кривизни заломлюючих поверхонь кришталика: радіус передньої поверхні може змінюватись від 10 до 5.5 мм, а задньої - від 6 до 5.5 мм. Відстань, на якій око розглядає предмет без напруги акомодації, називають відстанню найкращого зору. Типово ця відстань складає 25 см.
ока за рахунок зміни кривизни заломлюючих поверхонь кришталика: радіус передньої поверхні може змінюватись від 10 до 5.5 мм, а задньої - від 6 до 5.5 мм. Відстань, на якій око розглядає предмет без напруги акомодації, називають відстанню найкращого зору. Типово ця відстань складає 25 см.
Розглянемо недоліки оптичної системи ока. Властиві лінзам аберації для ока не є характерними, тому що: 1) сферична аберація усувається тим, що око пропускає лише центральний пучок світла; 2) хроматична аберація усувається тим, що око найбільш чутливе до вузької жовто-зеленої частини спектра; 3) астигматизм похилих променів усувається тим, що око автоматично встановлюється у напрямку предмета, що спостерігається, і до нього потрапляють лише центральні промені.
Оптичній системі ока притаманні такі недоліки: 1) Короткозорість (міопія) - подовжена форма очного яблучка, або занадто велика заломлююча здатність середовища ока при нормальній формі, що призводять до формування зображення перед сітківкою (рис. 6.45а). Усувається
міопія використанням розсіювальних лінз, тобто лінз з оптичною силою 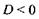 (рис. 6.45б).
(рис. 6.45б).
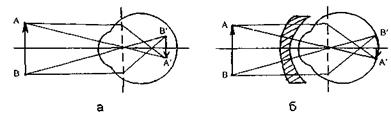
Рис. 6.45. Формування зображення в оці при міопії а та її усунення за допомогою розсіювальних лінз б.
2) Далекозорість (гіперметропія) - недостатня заломлююча здатність ока, внаслідок якої зображення формується позаду сітківки рис. 8.46а. Частково око усуває цей недолік за допомогою акомодації, але внаслідок границі акомодації застосовують збиральні лінзи, які посилюють заломлюючу здатність ока, тобто лінзи з оптичною силою 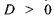 (рис. 6.46б).
(рис. 6.46б).
Рис. 6.46. Формування зображення при гіперметропії а та її усунення за допомогою збиральних лінз б.
3) Астигматизм, пов'язаний з порушенням правильної сферичної форми поверхні роговиці. Усувається циліндричними лінзами або лінзами спеціальної форми.
Оптичне зображення формується на сітківці. Відчуття світла з'являється при дії на світлочутливий елемент сітківки ока електромагнітного випромінювання з довжиною хвилі 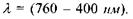 Сітківка має товщину
Сітківка має товщину 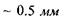 і складається з кількох шарів, що вміщують волокна зорового нерва (І), опорні утвори (II) і світлочутливі клітини (III), периферичні кінці яких мають різну форму (рис. 6.47). Подовжені кінці називають паличками, а конусоподібні -колбочками. На сітківці ока розташовано близько 130 мли паличок та 7 млн колбочок. Колбочки і палички розподілені нерівномірно: колбочки, головним чином, розташовані в центральній частині сітківки, тоді як палички - лише на краях сітківки.
і складається з кількох шарів, що вміщують волокна зорового нерва (І), опорні утвори (II) і світлочутливі клітини (III), периферичні кінці яких мають різну форму (рис. 6.47). Подовжені кінці називають паличками, а конусоподібні -колбочками. На сітківці ока розташовано близько 130 мли паличок та 7 млн колбочок. Колбочки і палички розподілені нерівномірно: колбочки, головним чином, розташовані в центральній частині сітківки, тоді як палички - лише на краях сітківки.
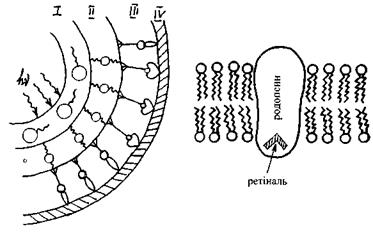
Рис. 6.47. Будова сітківки ока. Рис. 6.48. Родопсин, вбудований
в ліпідний бішар мембран зорових дисків.
Як палички, так і колбочки орієнтуються в напрямку світла своїми внутрішніми сегментами, які не вміщують зорових пігментів. Світлочутливі рецептори (палички і колбочки) межують з епітеліальним шаром (IV), який зменшує частку світла, що розсіюється і відбивається від задньої стінки ока.
Розглянемо будову і функції паличок. Зоровий пігмент паличок - родопсин - вбудований в ліпідний шар мембран так званих зорових дисків, що знаходяться в зовнішньому сегменті паличок (рис. 8.48). Родопсин - складний білок, що складається з білка опсину і хромофорної групи ретіналю ( 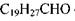 - альдегіду вітаміну А).
- альдегіду вітаміну А).
Ретіналь має кілька конформерів (циста трансконформери зображені на рис. 6.49). 11-цис-ретіналь точно вміщується в центр зв'язку молекули опсину.
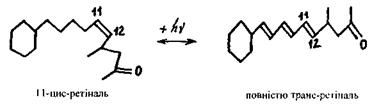
Рис. 6.49. Фотоізомеризація ретіналю.
Під дією падаючого випромінювання відбувається фотоізомеризація ретіналю. Внаслідок фотохімічного утворення трансформи стерична відповідність між опсином і ретіналем порушується, що супроводжується низкою конфор-маційних перебудов молекули опсину, які і призводять до виникнення нервового імпульсу (рис. 6.50).

Рис. 6.50. Механізм виникнення нервового імпульсу внаслідок фотоізомерізації ретіналю та конфірмаційної перебудови опсину.
Після освітлення палички коротким спалахом світла спостерігається ранній рецепторний потенціал (РРП), що триває близько 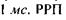 зумовлений переміщенням фіксованих на поверхні родопсину зарядів щодо подвійного шару ліпідних молекул. Величина РРП не перевищує
зумовлений переміщенням фіксованих на поверхні родопсину зарядів щодо подвійного шару ліпідних молекул. Величина РРП не перевищує 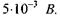 Через 1 мс спостерігається пізній рецепторний потенціал (ПРП), що зумовлений зміною проникності цитоплазматичних мембран зовнішніх сегментів паличок. При відсутності збудження мембрана має значну натрієву провідність. Під час збудження відбувається визволення із зорових дисків іонів калію і зачинення натрієвих каналів. Внаслідок цього значення мембранного потенціалу зсувається до значення калієвого рівноважного потенціалу. На відміну від інших, розглянутий рецепторний потенціал має не деполяризацій-ний, а гіперполяризаційний характер; мембранний потенціал рецепторної клітини під дією світла може змінюватись від
Через 1 мс спостерігається пізній рецепторний потенціал (ПРП), що зумовлений зміною проникності цитоплазматичних мембран зовнішніх сегментів паличок. При відсутності збудження мембрана має значну натрієву провідність. Під час збудження відбувається визволення із зорових дисків іонів калію і зачинення натрієвих каналів. Внаслідок цього значення мембранного потенціалу зсувається до значення калієвого рівноважного потенціалу. На відміну від інших, розглянутий рецепторний потенціал має не деполяризацій-ний, а гіперполяризаційний характер; мембранний потенціал рецепторної клітини під дією світла може змінюватись від 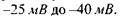
Палички мають високу світлочутливість, але не забезпечують розпізнавання кольорів. Саме тому їх називають апаратом ахроматичного (сутінкового) зору.
Колбочки мають нижчу світлочутливість, тобто сприймають світло при відносно великій освітленості сітківки, але при цьому забезпечують здатність розрізнення більш мілких деталей предмета і створюють відчуття кольору.
Чутливість ока до монохроматичного випромінювання різних довжин хвиль суттєво відрізняється. Для адаптованого до денного світла ока графік спектральної чутливості має вигляд кривої А з максимумом 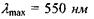 в жовто-зеленій ділянці спектра (рис. 6.51). При сутінковому зорі випромінювання різних довжин хвиль викликає однакове відчуття блакитно-сірого кольору. Максимум кривої спектральної чутливості (рис. 8.51, крива В) зсувається в бік менших довжин хвиль
в жовто-зеленій ділянці спектра (рис. 6.51). При сутінковому зорі випромінювання різних довжин хвиль викликає однакове відчуття блакитно-сірого кольору. Максимум кривої спектральної чутливості (рис. 8.51, крива В) зсувається в бік менших довжин хвиль 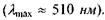 Доведено, що крива В співпадає з кривою поглинання родопсину. Що ж до кривої А, то її максимум на довжині хвилі
Доведено, що крива В співпадає з кривою поглинання родопсину. Що ж до кривої А, то її максимум на довжині хвилі 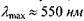 співпадає з максимумом спектра поглинання пігменту в найбільш поширеному типі колбочок.
співпадає з максимумом спектра поглинання пігменту в найбільш поширеному типі колбочок.
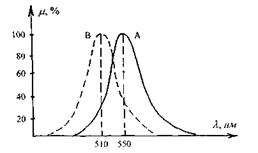
Рис. 6.51. Криві спектральної чутливості ока до денного А та сутінкового В світла.
Пігмент вміщує той самий 11-цисретіналь, що й родопсин. Що ж до білкової частини пігменту, то вона відрізняється і називається йодопсином. Поглинання світла йодопсинами призводить до появи РРП в колбочках.
Відповідно до теорії кольорового зору існує три типи колбочок, кожна з яких вміщує лише один тип йодопсину. Максимуми поглинання трьох типів йодопсинів відповідають 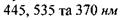 (рис. 6.52).
(рис. 6.52).
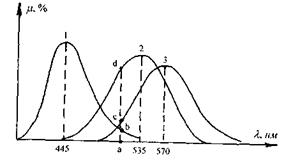
Рис. 6.52. Криві спектральної чутливості трьох типів йодопсинів: 1 - синьо-фіолетового; 2 - зеленого; 3 - червоного.
Рецептори всіх трьох типів збуджуються під дією світла будь-якої довжини хвилі, але у різних співвідношеннях, пропорційних ординатам відповідних кривих. Наприклад, під дією випромінювання з довжиною хвилі 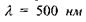 "червоні" рецептори збуджуються пропорційно ординаті ас, "зелені" -
"червоні" рецептори збуджуються пропорційно ординаті ас, "зелені" - 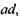 а "синьо-фіолетові" -
а "синьо-фіолетові" - 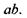 Імпульси від цих рецепторів передаються до головного мозку, де і утворюють єдине кольорове відчуття.
Імпульси від цих рецепторів передаються до головного мозку, де і утворюють єдине кольорове відчуття.
Якщо теорія сутінкового зору вважається нині досить обгрунтованою, про що свідчить присудження за її створення Нобелевської премії американському біофізику Дж. Уолду, то теорія кольорового зору перебуває в процесі постійного розвитку і вдосконалення. Так, одна з гіпотез про сприйняття кольорів базується на ідеї, що навіть окрема колбочка чи паличка може аналізувати весь спектральний склад видимого світла. Певні підстави для такої гіпотези є, а саме: зовнішні сегменти колбочок і паличок складаються з великої кількості (до 1500-1700) дисків, які утворюють
циліндричну стопу. Виходячи з теорії поширення світла в такій дисперсній системі, можна довести, що світло різної довжини хвилі послаблюється неоднаково при проходженні крізь стопу дисків кожної колбочки і палички. Підставою для цього є формула: 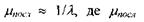 - коефіцієнт послаблення світлового потоку на одиницю довжини шляху променю, а
- коефіцієнт послаблення світлового потоку на одиницю довжини шляху променю, а 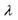 - довжина світлової хвилі. Звідси випливає, що синє світло поглинається в дисках сильніше, ніж червоне. Таким чином, при проходженні світла крізь стопу дисків із світлового потоку відбирається спочатку більш короткохвильова частина спектра, світловий потік поступово "червоніє", тобто в наступних дисках поглинається більш довгохвильова частина спектра. Це й дає змогу провести спектральний аналіз світлового променя, не використовуючи уяву про три типи йодопсинів.
- довжина світлової хвилі. Звідси випливає, що синє світло поглинається в дисках сильніше, ніж червоне. Таким чином, при проходженні світла крізь стопу дисків із світлового потоку відбирається спочатку більш короткохвильова частина спектра, світловий потік поступово "червоніє", тобто в наступних дисках поглинається більш довгохвильова частина спектра. Це й дає змогу провести спектральний аналіз світлового променя, не використовуючи уяву про три типи йодопсинів.
Дата добавления: 2020-11-18; просмотров: 836;
Поиск по сайту
Узнать еще
- ІV. Короткий зміст питань, що виносяться на державний іспит з теоретико – методичних основ фізичного виховання (основи теорії та методики спортивного тренування).
- Історія сучасних взаємовідносин людини й природного середовища. Екологія і практична діяльність людини. Життєвий простір. Екологічні основи раціонального природокористування.
- АКУСТИКА. ЕЛЕМЕНТИ ФІЗИКИ СЛУХУ. ОСНОВИ АУДІОМЕТРІЇ
- Змістовний модуль 3.Теоретичні основи математичної статистики
- Концептуальні основи створення системи неперервної освіти
- ЛЕКЦИЯ 11. ОСНОВИ ТЕОРИИ ВЫПОЛНЕНИЯ НАВИГАЦИОННЫХ МАНЕВРОВ
- Медико-біологічні основи фізичної культури

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
