Морфофункциональная организация спинного мозга
Спинной мозг – наиболее древний отдел ЦНС позвоночных. Он уже есть у ланцетника, наиболее примитивного представителя хордовых.
Спинной мозг – каудальный отдел ЦНС. Он помещается в позвоночном канале и имеет неодинаковую длину у разных представителей позвоночных.
У человека в каудальном отделе позвоночного канала собираются корешки каудальных отделов спинного мозга, образуя так называемый конский хвост.
Спинной мозг характеризуется сегментарным строением. В спинном мозге выделяют шейный, грудной, поясничный, крестцовый и копчиковый отделы. Каждый отдел состоит из нескольких сегментов. Шейный отдел включает 8 сегментов (С1 – С8), грудной – 12 (Th1 – Th12), поясничный – 5 (L1 – L5), крестцовый – 5 (S1 – S5) и копчиковый – 1-3 (Со1 – Co3). От каждого сегмента отходят две пары корешков, которые соответствуют одному из позвонков и покидают позвоночный канал через отверстие между ними.
Различают дорсальные (задние) и вентральные (передние) корешки. Дорсальные корешки образованы центральными аксонами первичных афферентных нейронов, тела которых лежат в спинномозговых ганглиях.
Вентральные корешки сформированы аксонами α- и γ-мотонейронов и немиелинизированными волокнами нейронов вегетативной нервной системы. Такое распределение афферентных и эфферентных волокон было установлено независимо друг от друга в начале XIX века Ч. Беллом (1811) и Ф. Мажанди (1822). Различное распределение функций в передних и задних корешках спинного мозга получило название закона Белла-Мажанди. Сегменты спинного мозга и позвонки соответствуют одному метамеру. Нервные волокна пары задних корешков идут не только к своему метамеру, но также выше и ниже – в соседние метамеры. Кожная область, в которой распределяются эти чувствительные волокна, называется дерматомом.
Количество волокон в дорсальном корешке гораздо больше, чем в вентральном.
Нейронные структуры спинного мозга.Центральную часть поперечного среза спинного мозга занимает серое вещество. Вокруг серого вещества располагается белое вещество. В сером веществе выделяют передние, задние и боковые рога, а в белом – столбы (вентральные, дорсальные, латеральные и т.д.).
Нейрональный состав спинного мозга достаточно разнообразен. Выделяют несколько видов нейронов. Тела нейронов спинномозговых ганглиев находятся вне спинного мозга. В спинной мозг входят аксоны этих нейронов. Нейроны спинномозговых ганглиев – это униполярные или псевдоуниполярные нейроны. В спинномозговых ганглиях лежат тела соматических афферентов, иннервирующих в основном скелетные мышцы. Тела других чувствительных нейронов находятся в ткани и в интрамуральных ганглиях автономной нервной системы и обеспечивают чувствительность только внутренних органов. Они бывают двух типов: крупные – диаметром 60-120 мкм и мелкие – диаметром 14-30 мкм. Крупные дают миелинизированные волокна, а мелкие – миелинизированные и немиелинизированные. Нервные волокна чувствительных клеток по скорости проведения и диаметру классифицируются на А-, В- и С-волокна. Толстые миелинизированные А-волокнадиаметром от 3 до 22 мкм и скоростью проведения от 12 до 120 м/с подразделяются на подгруппы: альфа-волокна – от мышечных рецепторов, бета-волокна – от тактильных и барорецепторов, дельта-волокна – от терморецепторов, механорецепторов и болевых рецепторов. К волокнам группы В относят миелинизированные волокна средней толщины со скоростью проведения возбуждения 3-14 м/с. По ним в основном передается ощущение боли. К афферентам типа С относят большинство безмиелиновых волокон толщиной не более 2 мкм и скоростью проведения до 2 м/с. Это волокна, которые идут от болевых, хемо- и некоторых механорецепторов.
В сером веществе спинного мозга выделяют следующие элементы:
1) эфферентные нейроны (мотонейроны);
2) вставочные нейроны;
3) нейроны восходящих трактов;
4) интраспинальные волокна чувствительных афферентных нейронов.
Моторные нейроны сосредоточены в передних рогах, где они образуют специфические ядра, все клетки которых посылают свои аксоны к определенной мышце. Каждое двигательное ядро тянется обычно на протяжении нескольких сегментов, поэтому и их аксоны, которые иннервируют одну и ту же мышцу, покидают спинной мозг в составе нескольких вентральных корешков.
В промежуточной зоне серого вещества локализуются вставочные нейроны. Их аксоны распространяются как внутри сегмента, так и в ближайшие соседние сегменты. Вставочные нейроны – разнородная группа, дендриты и аксоны которой не покидают пределов спинного мозга. Вставочные нейроны образуют синаптические контакты только с другими нейронами, и их большинство. На долю вставочных нейронов приходится около 97% от всех нейронов. По размерам они меньше α-мотонейронов, способны к высокочастотной импульсации (выше 1000 в сек.). Для проприоспинальных вставочных нейронов характерно свойство посылать длинные аксоны через несколько сегментов и оканчиваться на мотонейронах. Вместе с тем на эти клетки конвергируют волокна различных нисходящих трактов. Поэтому они являются релейными станциями на пути от вышележащих нейронов к мотонейронам. Особую группу вставочных нейронов образуют тормозные нейроны. К ним относятся, например, клетки Реншоу.
Нейроны восходящих трактов также целиком находятся в пределах ЦНС. Тела этих нейронов расположены в сером веществе спинного мозга.
Центральные окончания первичных афферентов имеют свои особенности. После вступления в спинной мозг афферентное волокно, как правило, дает начало восходящей и нисходящей ветвям, которые могут идти на значительные расстояния по спинному мозгу. Концевые разветвления одного нервного афферентного волокна имеют многочисленные синапсы на одном мотонейроне. Кроме того, установлено, что одно волокно, идущее от рецептора растяжения, образует синапсы почти со всеми мотонейронами данной мышцы.
В дорсальной части дорсального рога располагается желатинозная субстанция Роланда.
Наиболее точное представление о топографии нервных клеток серого вещества спинного мозга дает разделение его на последовательные слои или пластины, в каждой из которых группируются, как правило, однотипные нейроны.
Согласно этим данным, все серое вещество спинного мозга было разделено на 10 пластин (Рексед) (рис. 2.2).
I – краевые нейроны – дают начало спиноталамическому тракту;
II-III – желатинозная субстанция;
I-IV – в целом первичная сенсорная область спинного мозга (афферентация от экстерорецепторов, афферентация от рецепторов кожной и болевой чувствительности);
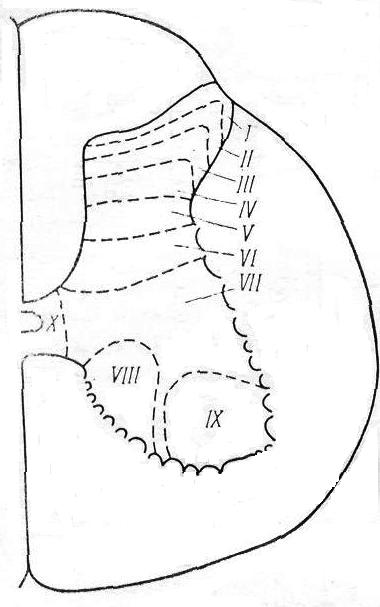
Рис. 2.2. Деление серого вещества спинного мозга на пластины (по Рекседу)
V-VI – локализуются вставочные нейроны, которые получают входы от задних корешков и нисходящих путей (кортико-спинальный, рубро-спинальный);
VII-VIII – располагаются проприоспинальные вставочные нейроны (от проприорецепторов, волокон вестибуло-спинального и ретикуло-спи-
нального трактов), аксоны проприоспинальных нейронов;
IX – содержит тела α- и γ-мотонейронов, пресинаптические волокна первичных афферентов от мышечных рецепторов растяжения, окончания волокон нисходящих трактов;
X – окружает спинномозговой канал и содержит наряду с нейронами значительное количество глиальных клеток и комиссуральных волокон.
Свойства нервных элементов спинного мозга. Спинной мозг человека содержит примерно 13 миллионов нейронов.
α-мотонейроны – крупные клетки с длинными дендритами, имеющие до 20 000 синапсов, большая часть которых образована окончаниями интраспинальных вставочных нейронов. Скорость проведения по их аксону составляет 70-120 м/с. Характерны ритмические разряды с частотой не выше 10-20 имп/с, что связано с выраженной следовой гиперполяризацией. Это выходные нейроны. Они осуществляют передачу сигналов скелетным мышечным волокнам, выработанных в спинном мозге.
γ-мотонейроны – более мелкие клетки. Диаметр их не более 30-40 мкм, они не имеют непосредственного контакта с первичными афферентами.
γ-мотонейроны иннервируют интрафузальные (внутриверетенные) мышечные волокна.
Они моносинаптически активируются волокнами нисходящих трактов, что играет важную роль в α-, γ-взаимодействии. Скорость проведения по их аксону ниже – 10-40 м/с. Частота импульсов выше, чем у α-мото-
нейронов, – 300-500 имп/с.
В боковых и передних рогах находятся преганглионарные нейроны вегетативной нервной системы – аксоны их направляются к клеткам ганглиев симпатической нервной цепочки и к интрамуральным ганглиям внутренних органов.
Тела симпатических нейронов, аксоны которых образуют преганглионарные волокна, располагаются в интермедиолатеральном ядре спинного мозга. Их аксоны относятся к группе В-волокон. Им свойственна постоянная тоническая импульсация. Одни из этих волокон участвуют в поддержании сосудистого тонуса, а другие обеспечивают регуляцию висцеральных эффекторных структур (гладкой мускулатуры пищеварительной системы, железистых клеток).
Тела парасимпатических нейронов образуют крестцовые парасимпатические ядра. Они располагаются в сером веществе крестцового отдела спинного мозга. Для многих из них характерна фоновая импульсная активность, частота которой возрастает, например, по мере повышения давления в мочевом пузыре.
Дата добавления: 2016-07-27; просмотров: 3267;
Поиск по сайту
Узнать еще
- I. Синусы твердой оболочки головного мозга.
- II. Анатомия продолговатого мозга, ретикулярная формация
- II. Организация дезинфекционных и стерилизационных мероприятий в организациях, осуществляющих медицинскую деятельность
- II. Функционально-структурная организация и программное обеспечение персонального компьютера
- IV. проводящие пути головного и спинного мозга
- V Патопсихологическое – при нарушении целостности мозга происходит нарушение психической деятельности
- XXII. ОРГАНИЗАЦИЯ РАБОТ ПО ОБЕСПЫЛИВАНИЮ РУДНИЧНОГО ВОЗДУХА
- А Нейропсихологические синдромы поражения задних отделов коры больших полушарий головного мозга.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
