Особенности и методы изучения докембрийских пород.
С позиций развития органической жизни геологическую стадию развития планеты делят на два этапа – криптозой (этап скрытой жизни) и фанерозой (этап явной жизни). Первый охватывает докембрийский этап, второй – всё от кембрия до наших дней. Докембрий представляет около 85% времени геологической истории Земли, однако изучен он намного слабее, чем фанерозой. Это обусловлено с глубокими изменениями докембрийских пород, подвергшихся метаморфизму, складчатости и гранитизации. Между тем, выходы докембрийских пород на дневную поверхность составляют до 20% площади континентального блока и с ними связаны многие полезные ископаемые.
Первоначально образования докембрия имеют как осадочное, так и магматическое происхождение. Затем они в большей или меньшей степени подвергались метаморфизму, основной вид которого в докембрии – региональный. Осадочные породы при глубоком метаморфизме превратились в гнейсы, кварциты, кристаллические сланцы, мраморы, при меньшем – в серицит-хлоритовые сланцы, филлиты, кварцитовидные песчаники, мраморовидные известняки другие образования. Вулканогенные породы основного состава переходят в амфиболиты и амфиболовые гнейсы при сильном метаморфизме, при слабом изменения незначительны. Кислые и средние породы при сильном метаморфизме дают лептиты. Выделяют ортопороды, образовавшиеся по магматитам, и парапороды по осадочным отложениям.
Степень метаморфизма докембрийских пород различна и убывает обычно снизу вверх. Выделяют три главных фации метаморфизма – зелёных сланцев, амфиболитовую и гранулитовую, отражающих разные термодинамические обстановки. Особо выделяется фация глаукофановых сланцев, которая является разновидностью амфиболитовой. Фация зеленых сланцев характеризуется условиями низких температур и давления, типичных для небольших глубин, когда образуются такие минералы как хлорит, серицит, эпидот, актинолит.
Для амфиболитовой фации характерны большие давления, высокие температуры и наличие воды, для глаукофановой – всё то же самое, но низкая температура. Эта фация обычна для зон разломов. Глаукофан – Na-Al амфибол. К породам амфиболитовой фации относится основной объём метаморфических пород – гнейсы, амфиболиты, кристаллические сланцы, лептиты. Гранулитовая фация характеризует условия ещё больших давлений и температур, но без воды. Гранулиты – это гиперстен-гранатовые гнейсы без слюд и амфиболов.
В докембрии широко развиты процессы гранитизации с формированием мигматитов (тонкое переслаивание гранитов и гнейсов), а сам процесс называется мигматизацией. При более глубокой переработке большие объёмы гнейсов, кристаллических сланцев, вулканогенных пород в условиях больших температур и давлений и под влиянием горячих растворов, с привносом ряда элементов и соединений, превращаются в метасоматические граниты (процессы анатексиса и палингенеза). В итоге возникли огромные массивы гранитов, слагающие обширные площади докембрийских платформ. Гранитизация очень затрудняет установление первичного состава пород.
Докембрийские образования обычно сильно дислоцированы, их отличает наличие сложных складок многих порядков. По тектоническим признакам в докембрийских разрезах возможно выделение ряда структурных этажей, свидетельствующих о проявлении ряда эпох складчатости. Исследователям приходится мириться с приблизительностью и неточностью расчленения и корреляции докембрийских образований по степени метаморфизма, глубине магматической и тектонической переработки, петрографическим особенностям пород. Палеонтологический метод практически не работает для стратификации архейских и нижнепротерозойских образований, поэтому для расчленения докембрия используются минералого-петрографические, геофизические, структурно-тектонические, а также радиогеохронологические методы исследований. Отложения позднего докембрия можно расчленять с помощью палеонтологического метода, поскольку в них имеются органические остатки. Геофизические методы обычно предваряют геологические.
Для изучения докембрия широко используется метод выделения конкретных структурно-вещественных комплексов пород, отличающихся друг от друга по составу, условиям залегания, степени метаморфизма. Выясняют взаимоотношения этих комплексов, порядок залегания, условия образования. При этом большое значение имеют петрофизические методы. Для определения возраста пород используются радиогеохронологические методы. Однако следует помнить об ограничениях, поскольку мы имеем дело не с возрастом первичных пород, а минералами в них, образовавшихся, как правило, при метаморфизме.
Стратиграфия
В настоящее время общепринятой шкалы расчленения докембрия нет. Имеется международная шкала и шкала Северной Евразии (табл. 6.1), где представлены крупные стратиграфические единицы, границы между которыми совпадают с проявлениями диастрофизма. В 1872 году американский геолог Дж. Дэна наиболее древние метаморфические образования назвал архейскими (греч. «археос» – древний), а затем В. Эммонс в 1888 г. выделил верхнюю часть древнейших толщ под названием протерозой («протерос» – первичный, «зоэ» жизнь). Эти подразделения долгое время были в ранге эр (групп), но из-за большой продолжительности (около 2 млрд лет каждое) по сравнению с эрами фанерозоя, потребовались более крупные геохроны (стратоны).
Таблица 6.1
Международная шкала Шкала Северной Евразии
Млн лет
| Эон | Эра | Период | Акротема | Эонотема | Эратема | Система | |
| Основание кембрия 540 | Основание кембрия 540 | ||||||
| Протерозой | Неопротерозой | Протерозой | Верхний р и ф е й | Венд 650±20 | Верхний нижний | ||
| Мезопротерозой | Верхний (каратавий) | 1000±50 | |||||
| Палеопротерозой | Средний (юрматиний) | 1350±50 | |||||
| Архей | Неоархей | Нижний (бурзяний) | 1650±50 | ||||
| Мезоархей | нижний | Верхний карелий | 1900±50 | ||||
| Палеоархей | Нижний карелий | 2500±50 | |||||
| Архей | верхний | 3150±50 | |||||
| нижний | 4000±50 |
В ныне действующем стратиграфическом кодексе архей (4,0-2,6 млрд лет) и протерозой (2,6-0,65 млрд лет) имеют ранг акронов (акротем), делящихся каждый на два эона – ранний и поздний, которые в стратиграфической шкале соответствуют эонотемам – нижней и верхней. Нижне – и верхнеархейская эонотемы не имеют пока более дробных подразделений в стратиграфической шкале, а нижний протерозой делится на две эратемы – нижнюю и верхнюю. В России их называют нижний и верхний карелий, поскольку наиболее представительные и хорошо изученные разрезы протерозоя находятся в Карелии. Верхний протерозой разделяется на рифей и венд. Первое наименование принято по древнему названию Уральских гор, второе – по названию древнего племени вендов, живших на территории Прибалтики. Ранг рифея – эонотема, разделяющаяся на три эратемы – нижне-, средне- и верхнерифейские. Венд – это скорее период (система).
В западной литературе принято трёхчленное деление архея и протерозоя. Выделяются палео-(4,0-3,5 млрд лет), мезо-(3,5-3,0 млрд лет), неоархей (3,0-2,5 млрд лет) и палео-(2,5-1,6 млрд лет), мезо-(1,6-1,0 млрд лет), неопротерозой (1,0-0,65 млрд лет). Вместе с тем, описывая события докембрийского этапа длительностью свыше 3,5 млрд лет, следует помнить, о некоторой условности и спорности расчленения докембрийских отложений. В этот громадный промежуток времени не существовало таких групп организмов, которые испытывали бы быстрое развитие, что является непременным условием дробной стратиграфической шкалы – основы расчленения отложений фанерозоя.
Органический мир
Жизнь зародилась в раннем архее и была первоначально представлена прокариотами – одноклеточными организмами, не имевшими обособленного ядра. Они прошли сложный путь развития и к середине архея существовали две самостоятельные ветви – бактерии и цианобионты (синезеленые водоросли). Установлены также узорчатые неслоистые карбонатные образования – катаграфии, грибы.
В мелких ваннах, сильно прогретых солнцем, в бескислородной среде с преобладанием углекислоты, в условиях парникового эффекта, в своеобразном абиогенном «бульоне», в окружении фумарол и вулканов, могли возникнуть высокополимерные нуклеиновые кислоты, вернее их спиральные нити, обладающие способностью синтезировать себе подобные образования. Первоначально возникли коацерватные капли, обособленные от внешней среды полупроницаемыми мембранами, через которые был возможен обмен веществ с окружающей средой. Затем образовались протобионты, потом предбиологические многомолекулярные системы и дезоксирибонуклеиновые кислоты (ДНК) – основной материал для формирования клеток. Существует также парадигма панспермии, предполагающая занесение микроскопических зародышей на Землю. При изучении метеорита Мерчисон из Австралии обнаружено 18 аминокислот, 6 из которых входят в состав белков. Еще в одном метеорите обнаружено углистое вещество, представляющее продукты жизнедеятельности спор и бактерий.
Именно в то время, когда появилась возможность копирования белков, т.е. возникли первые примитивные клетки, очевидно и была преодолена граница между неживым и живым мирами. Первыми живыми организмами являлись бактерии, превращавшие неорганические соединения в органические, используя солнечный свет. Бактерии поглощали сероводород, выделяя при этом серу. И только синезеленые водоросли разлагали воду, продуцируя кислород. Среди организмов были автотрофные, использующие неорганическое вещество, и гетеротрофные, питающиеся автотрофами, в том числе сапрофаги, существующие за счёт разлагающихся органических остатков.
Продукты жизнедеятельности синезеленых водорослей – строматолитов и бактерий – онколитов встречены в отложениях возрастом 3,7 млрд лет. Строматолиты – это скорлуповатые карбонатные корочки, нарастающие друг на друга и слагающие иногда крупные рифоподобные постройки мощностью в десятки и сотни метров и протяжённостью в сотни километров. Онколиты – образования из карбонатных желваков концентрически-скорлуповатой текстуры. Все они жили в бескислородной мелководной среде, а слои воды толщиной более 10 м защищали их от губительного ультрафиолетового излучения.
В раннем протерозое появились первые многоклеточные (простейшие губки, кишечнополостные). Деятельность синезеленых водорослей привела к появлению кислорода в атмосфере и гидросфере. Содержание кислорода по А.Ю. Розанову (1986) достигло точки Юри (0,1% современного уровня) 3 млрд лет назад, точки Пастера (1%) – 2,2-2,5 млрд лет назад, Беркнера-Маршала (10%) – 1,8-2,0 млрд лет. Появление свободного кислорода при достижении точки Пастера позволило организмам перейти от энергии анаэробного брожения к более выгодному (в 30-50 раз) окислению при дыхании. Появление в верхних слоях атмосферы озонового слоя, защищающего биос от гибели, отвечает 10% от современного содержания кислорода. Нарастание фитомассы определило повышение содержаний кислорода, а отсюда уменьшение подвижности железа. Следствием этого является исчезновение к рифею железистых кварцитов и появление красноцветных формаций. Одновременно продолжалось изъятие углерода. Это фиксируется резким в истории Земли приростом глобального обилия и разнообразия строматолитов. В процессе развития органической жизни в значительных объемах наряду с накоп-
лением в атмосфере этого элемента происходило его изъятие из неё. Если на рубеже архея и про-

терозоя она была преимущественно азотной, то нарастание количеств О2 происходило одновременно с изъятием из нее части азота.
В позднем протерозое отмечается три этапа развития органической жизни. Особенно пышное развитие получили синезеленые водоросли, остатки жизнедеятельности которых – строматолиты имеют определяющее значение для стратиграфии рифея (рис. 6.1). Именно по ним выделены стратиграфические подразделения рифея, и впервые у нас в стране. Первый этап связан с началом позднего протерозоя, когда появились эукариоты – организмы, клетки которых имели обособленные ядра. Эта биота перешла на кислородное дыхание, и среди неё появились первые планктонные организмы. Второй этап развития организмов начался в среднем рифее, когда содержание кислорода достигло более 10%, что обеспечило их защиту только однометровым слоем воды. Появились примитивные многоклеточные организмы среди растений и животных, причем не только неподвижные, но и бентосные формы, а также подвижные илоеды, следы жизнедеятельности которых представляют катаграфии (1.2 млрд лет). Последние представляют собой желвачки из слипшихся комочков и пузырьков. Отмечаются также акритархи – остатки зоо- и фитопланктона и грибы. Третий (вендский) этап характеризуется становлением основных типов животного мира, в первую очередь, многоклеточных.
Вендская фауна и флора (рис. 6.2) отличалась большим разнообразием и богатством многоклеточных и одноклеточных форм. Фауна представлена примитивными кишечнополостными (медузы, полипы, морские перья), червями, животными, близкими к членистоногим и иглокожим, флора только водорослями. Отличительной особенностью вендских форм является отсутствие минерального скелета. Однако покровы их были уплотнены и поэтому на мягких грунтах остались отпечатки тел животных. Отмечаются многочисленные следы жизнедеятельности вендской фауны. Наиболее многочисленные формы найдены в Австралии на руднике Эдиакар (эдиакарская фауна), на берегу Белого моря, в Южной Африке. До 70% находок – кишечнополостные, представленные медузоидными формами до 0,5 м в диаметре, одиночными и колониальными полипами. В венде зародились древнейшие многоклеточные животные с хитиновой оболочкой – прообразом минерального скелета.
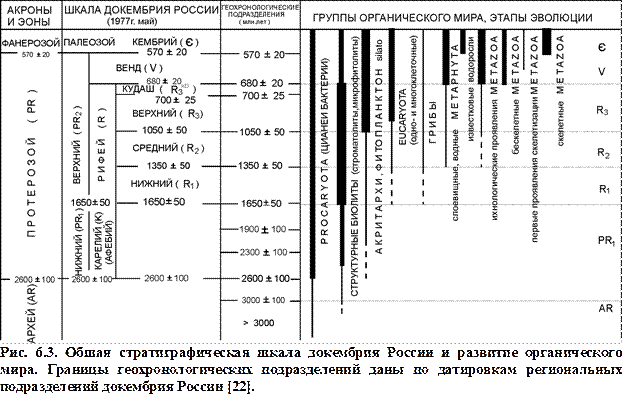 Таким образом, в течение докембрия появились и эволюционировали представители как растительного, так и животного царства (рис. 6.3). Вместе с тем наибольшее распространение группы органического мира получили в конце позднего протерозоя.
Таким образом, в течение докембрия появились и эволюционировали представители как растительного, так и животного царства (рис. 6.3). Вместе с тем наибольшее распространение группы органического мира получили в конце позднего протерозоя.
| <== предыдущая лекция | | | следующая лекция ==> |
| | | А) Актуалізація даних |
Дата добавления: 2020-10-25; просмотров: 333;
Поиск по сайту
Узнать еще
- I. Гидрометаллургические методы
- I. Погрешности механической обработки. Точность обработки. Методы их расчёта
- I. Специфические особенности процесса воспитания в сравнении с процессом обучения.
- II. Методы исследования истории медицины.
- II. Пирометаллургические методы.
- II. Урок изучения нового материала.
- II.1.5. ОСОБЕННОСТИ РАЗВИТИЯ ЛИЧНОСТИ И ЭМОЦИОНАЛЬНО-ВОЛЕВОЙ СФЕРЫ
- II.II. Репродуктивные методы.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по истории
