Ферменты, не подчиняющиеся кинетике Михаэлиса-Ментен
Ферменты, кинетика которых не подчиняется гиперболическому закону Михаэлиса-Ментен, являются аллостерическими ферментами. Аллостерические ферменты отличаются от обычных ферментов по ряду признаков:
1) имеют олигомерную структуру, т.е. обладают 4-й структурой и состоят из субъединиц, объединенных в единое целое связями слабого характера;
2) наряду с активными центрами имеют специальные регуляторные (или аллостерические) центры. Активные и аллостерические центры пространственно разобщены. К аллостерическим центрам могут присоединяться низкомолекулярные соединения, называемы эффекторами или модуляторами. В качестве эффекторов могут выступать промежуточные метаболиты или конечные продукты метаболических путей;
4) молекула эффектора, связываясь в аллостерическом центре, вызывает изменение конформации фермента. Это приводит в свою очередь к изменению конформации активного центра, в результате активность фермента увеличивается (активация) либо уменьшается (ингибирование). Эффекторы обратимо взаимодействуют с аллостерическими центрами. Они не обладают явным сходством с субстратами, а фермент проявляет исключительно высокую специфичность к молекуле эффектора;
5) у аллостерических ферментов наблюдается отклонение зависимости скорости ферментативной реакции (v) от концентрации субстрата (S) или аллостерического эффектора (Э), а именно отклонение от простых кинетических закономерностей типа гиперболического закона Михаэлиса-Ментен.
Термин аллостерический был предложен Ж. Мано и Ф. Жакобом в 1961 г. Они предположили, что взаимодействие эффектора с аллостерическим центром вызывает в молекуле фермента конформационные изменения, затрагивающие активный центр, в результате чего активность фермента изменяется.
Не все ферменты в клетке являются аллостерическими, но многие аллостерические ферменты являются ключевыми ферментами, определяющими скорость лимитирующих стадий обменных процессов в клетке.
Для аллостерических ферментов характерным свойством является наличие кооперативных эффектов: присоединение первой молекулы соответствующего лиганда (субстрата к активному центру или эффектора к аллостерическому центру) сопровождается конформационными изменениями, которые изменяют его сродство к субстрату или эффектору.
Кооперативные эффекты подразделяют на гомотропные и гетеротропные. Гомотропные эффекты, при которых взаимодействия с лигандами могут быть кооперативными и антикооперативными, наблюдаются для идентичных лигандов, например, для молекул субстрата (а также для молекул кофермента или ингибитора). Гетеротропные эффекты, при которых взаимодействия, также являющиеся либо кооперативными, либо антикооперативными, наблюдаются для молекул различных лигандов.
Кооперативные эффекты, кроме того, подразделяют на положительные и отрицательные (антикооперативные). В случае положительной кооперативности присоединение первой молекулы лиганда вызывает конформационные изменения в белковой молекулы, которые передаются и облегчают присоединение последующих молекул лиганда (при этом зависимость скорости реакции от концентрации лиганда является S-образной) (рис. 2.4.1).

Рис. 2.4.1. Зависимость скорости реакции от концентрации субстрата
При антикооперативности присоединение первой молекулы лиганда вызывает такие конформационные изменения в белке, которые затрудняют присоединение последующих молекул лиганда к соответствующим центрам (зависимость скорости реакции от концентрации лиганда является гиперболической, но это не равнобочная гипербола Михаэлиса-Ментен) (рис. 2.4.1).
Такие различия легко увидеть, если линеаризовать зависимости скорости реакции от концентрации лигандов для трех случаев: положительной и отрицательной кооперативности и кинетики Михаэлиса-Ментен (рис.2.4.2).
При построении графика в координатах двойных обратных величин S-образная кривая преобразуется в кривую выпуклую к оси абсцисс. В последние годы было показано, что отдельные ферменты обнаруживают гиперболическую зависимость скорости от концентрации субстрата. Однако форма гиперболы в этих случаях отличается от классической гиперболы Михаэлиса-Ментен: в координатах Лайнуивера-Берка последняя преобразуется не в прямую линию, а в кривую выпуклую к оси ординат

Рис. 2.4.2. Линеаризация кривых, приведенных на рис. 2.4.1
У аллостерических ферментов, так же как и у нерегуляторных ферментов, наблюдается «насыщение» субстратом (рис. 2.4.3).
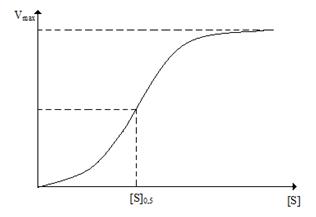
Рис. 2.4.3. Кривая с насыщением для аллостерического фермента
Хотя на сигмоидной кривой насыщения субстратом для аллостерических ферментов можно найти точку, в которой скорость реакции равна половине от максимальной скорости, эта величина не соответствует величине Km, поскольку поведение аллостерических ферментов не описывается гиперболической зависимостью, вытекающей и уравнения Михаэлиса-Ментен.
Первым белком с олигомерной структурой, проявляющим кооперативные эффекты при связывании кислорода, был гемоглобин. Кооперативность у гемоглобина была установлена Бором задолго до обнаружения подобных свойств у какого-либо фермента. Выявлению кооперативных свойств гемоглобина при его функционировании помогло сравнение с миоглобином, белком, не проявляющим кооперативных свойств и служащим для запасания кислорода в мышцах.
Впервые кооперативные эффекты были описаны Хиллом при анализе кривой насыщения гемоглобина кислородом (рис.2.4.4).
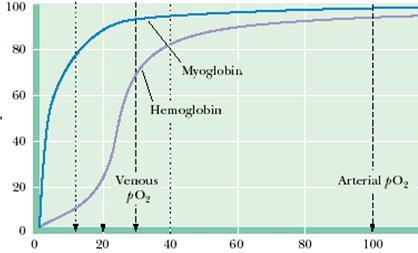
Рис.2.4.4. Кривые насыщения миоглобина и гемоглобина кислородом
Уравнение для описания кооперативных взаимодействий при связывании гемоглобина кислородом было предложено Хиллом.
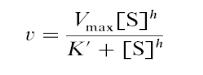
Где, v – наблюдаемая скорость ферментативной реакции;
VMAX – максимальная скорость реакции при концентрации субстрата, стремящейся к бесконечности;
К′– концентрация субстрата, при которой скорость равна половине от максимальной. Поскольку К′ не соответствует величине Км в данном случае вместо символа Км используют символы ([S]0,5, К0,5);
h – суммарное число связывающих центров в олигомерной молекуле фермента или коэффициент Хилла, или коэффициент кооперативных взаимодействий. Является мерой силы кооперативных взаимодействий при связывании лиганда. Коэффициент Хилла должен быть целым числом, но экспериментально определяемые величины часто не являются таковыми. В этих случаях за число связывающих участков принимают следующее, большее по величине целое число.
Например, экспериментально определенная величина h составляет 1,65. Это говорит о наличии минимум двух связывающих сайтов и что сайты проявляют уровень кооперативности выше среднего. Однако нельзя сказать, что фермент имеет только два связывающих центра. У него может быть три или 4 или более связывающих центров со слабо выраженной кооперативностью. Поэтому величина два в этом примере говорит о минимальном числе возможных связывающих центров.
Дата добавления: 2020-10-01; просмотров: 784;
Поиск по сайту
Узнать еще
- Анализ уравнения Михаэлиса-Ментен
- Дисперсные системы, подчиняющиеся уравнению Рэлея.
- Коферменты, простетические группы, ионы металлов
- Препараты на основе очищенных продуктов метаболизма микроорганизмов (витамины, аминокислоты, ферменты, антибиотики и др.)
- Уравнение Михаэлиса-Ментен
- Уравнения переноса в физико-химической кинетике

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
