Поступление вещества в клетки
Благодаря содержанию растворов солей, Сахаров и других осмотически активных веществ, клетки характеризуются наличием в них определенного осмотического давления. Например, давление в клетках животных (морских и океанических форм) достигает 30 атм и более. В клетках растений осмотическое давление является еще большим. Разность концентрации веществ внутри и снаружи клетки называют градиентом концентрации.
Поступление веществ в клетки животных, равно как и удаление их из клеток, связано с проницаемостью клеточной мембраны для молекул или ионов, а также со свойствами веществ. Клеточная мембрана регулирует обмен различными веществами между клеткой и средой. Поддержание мембраны и ее проницаемость обеспечиваются клеточной энергией.
Известно несколько путей поступления веществ в клетки. В частности, различают пассивный, катализируемый и активный транспорт веществ в клетки, а также проникновение веществ в клетки путем эндоцитоза в виде фагоцитоза и пиноцитоза. Пассивный, катализируемый и активный транспорт обеспечивают проникновение в клетки лишь малых молекул, тогда как эндоцитоз ответственен за поступление в клетки макромолекул (белков, по-линуклеотидов, полисахаридов) и разных твердых частиц, включая бактерии. В то же время клетки способны секретировать различные вещества в окружающую их среду. Этот процесс называют экзоцитозом.
Пассивный транспорт веществ в клетки обеспечивается диффузией через мембрану по градиенту концентрации. Молекулы обычно переходят из области высокой концентрации в область более низкой концентрации.
Количество работы, затрачиваемой на обеспечение транспорта молекул в клетку против градиента концентрации, можно определить, исходя из допущенияпростой реакции, в которой Ац есть концентрация молекул за пределами клетки, a Ai есть концентрация молекул внутри клетки. Эту реакцию можно описать константой равновесия в виде уравнения:
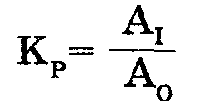
Между тем константа равновесия Кр связана со свободной энергией реакции отношением в виде G = RTInКр, где R есть около 2 кал/моль, а Т есть 25°С (температура, при которой протекают многие биологические реакции). Допуская, что совместная энергия гидролиза АТФ к АДФ обеспечивает эту реакцию с 50-процентной эффективностью, можно далее допустить, что транспортная система будет располагать примерно 3500 калориями (из общего количества энергии в 7000 калорий) на 1 моль АТФ, гидролизуемого при определенных физиологических условиях. Следовательно, константа равновесия будет равна:
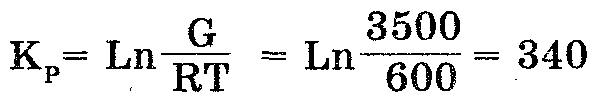
Важнейший вывод их этих заключений состоит в том, что работа, необходимая для транспорта какой-либо молекулы, не зависит от абсолютных концентраций. Она зависит от отношений между концентрациями внутри и вне клетки.
Когда транспортируются незаряженные молекулы, то пассивный транспорт определяется только градиентом концентрации, т. е. разностью концентрации вещества на разных сторонах мембраны. Если же молекулы транспортируемого вещества заряжены, то к влиянию градиента концентрации добавляется влияние электичес-ких потенциалов по обе стороны мембраны. Градиент концентрации и электрический градиент в совокупности составляют электрохимический потенциал, который позволяет транспорт в клетку только положительно заряженных ионов.
Можно сказать, что пассивный транспорт веществ в клетки осуществляется обычной диффузией через клеточную мембрану, причем скорость диффузии вещества зависит от его растворимости в мембране, коэффициента диффузии в мембране и от разности концентрации веществ в клетке и за ее пределами (в среде). Этим путем в клетку проникают вода, двуокись углерода и молекулы органических веществ, способные хорошо растворяться в жирах. Вещества в клетку проникают через поры, имеющиеся в клеточной мембране. Пассивный транспорт не зависит от энергии, обеспечиваемой АТФ.
Известна катализируемая, или так называемая «облегченная» диффузия, при которой скорость диффузии разных веществ, например, Сахаров, аминокислот и нуклеозидов через мембрану повышается с помощью белков (ферментов). Как и обычная диффузия, «облегченная» диффузия тоже зависит от градиента концентрации, однако здесь имеются подвижные «переносчики», роль которых выполняют ферменты. Находясь в составе мембраны, ферменты действуют в качестве «переносчиков» молекул веществ, проникая (диффундируя) на противоположную сторону мембраны, где они освобождаются от переносимых веществ. Поскольку «облегченная» диффузия веществ является переносом по градиенту концентрации, она тоже непосредственно не зависит от энергии, обеспечиваемой АТФ.
Активный транспорт веществ в клетку отличается от пассивного (диффузии) тем, что вещество переносится против градиента концентрации, т. е. из области низкой концентрации в область более высокой концентрации. Активный транспорт связан со способностью мембраны поддерживать разность электрических потенциалов (помимо поддержания разности в концентрациях веществ внутри и снаружи клетки), под которыми понимают различия между электрическими потенциалами внутри и вне клетки, а также с затратами энергии на работу в виде перемещения веществ против электрохимического градиента, т. е. «вверх».
Энергия для транспорта обеспечивается фосфоэнолпируватом, фосфатная группа которого и часть химической энергии которого передаются белкам, часть которых используется всеми сахарами, транспортируемыми фосфотрансферазной системой, а часть специфична для отдельных Сахаров. Конечный белок содержится в мембране и ответственен за транспорт и фосфорилирование Сахаров.
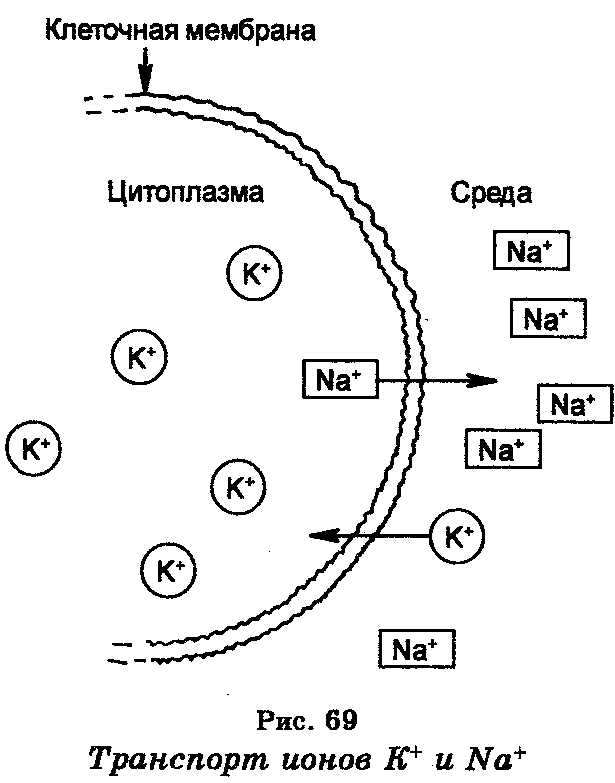 Активный транспорт особенно эффективен в случае переноса ионов. Реакции, обеспечивающие активный транспорт, происходят в мембране и сопряжены с реакциями, дающими свободную энергию. Ферменты, катализирующие эти реакции, также локализованы в мембране. Примером активного транспорта веществ является транспорт ионов натрия и калия (рис. 69), который определяет клеточный мембранный потенциал. Концентрация ионов натрия (Na+) внутри большинства клеток является меньшей, чем в среде, тогда как концентрация ионов калия (К+) внутри клеток является в 10—20 раз большей, чем в среде. В результате этого ионы Nа+ стремятся проникнуть из среды в клетку, а ионы К+, наоборот, выйти из клетки в среду. Поддержание концентрации этих ионов в клетке и в окружающей среде обеспечивается благодаря наличию в клеточной мембране системы, которая является ионным «насосом» и которая откачивает ионы Na+ из клетки в среду и накачивает ионы К+ в клетку из среды. Работа этой системы, т.е. движение ионов против электрохимического градиента, обеспечивается энергией, которая генерируется гидролизом АТФ, причем фермент АТФ-аза, катализирующий эту реакцию, содержится в самой мембране и, как считают, выполняет роль натриево-калиевого «насоса», генерирующего мембранный потенциал. Энергия, освобождаемая при гидролизе одной молекулы АТФ, обеспечивает транспорт за пределы клетки трех ионов Na+ и внутрь клетки двух ионов К+.
Активный транспорт особенно эффективен в случае переноса ионов. Реакции, обеспечивающие активный транспорт, происходят в мембране и сопряжены с реакциями, дающими свободную энергию. Ферменты, катализирующие эти реакции, также локализованы в мембране. Примером активного транспорта веществ является транспорт ионов натрия и калия (рис. 69), который определяет клеточный мембранный потенциал. Концентрация ионов натрия (Na+) внутри большинства клеток является меньшей, чем в среде, тогда как концентрация ионов калия (К+) внутри клеток является в 10—20 раз большей, чем в среде. В результате этого ионы Nа+ стремятся проникнуть из среды в клетку, а ионы К+, наоборот, выйти из клетки в среду. Поддержание концентрации этих ионов в клетке и в окружающей среде обеспечивается благодаря наличию в клеточной мембране системы, которая является ионным «насосом» и которая откачивает ионы Na+ из клетки в среду и накачивает ионы К+ в клетку из среды. Работа этой системы, т.е. движение ионов против электрохимического градиента, обеспечивается энергией, которая генерируется гидролизом АТФ, причем фермент АТФ-аза, катализирующий эту реакцию, содержится в самой мембране и, как считают, выполняет роль натриево-калиевого «насоса», генерирующего мембранный потенциал. Энергия, освобождаемая при гидролизе одной молекулы АТФ, обеспечивает транспорт за пределы клетки трех ионов Na+ и внутрь клетки двух ионов К+.
Система Na+ + К+ —АТФ-аза помогает поддерживать ассиметрическое распределение ионов калия при высокой концентрации последнего в клетках. Ионы калия участвуют в регуляции многих клеточных функций, включая поток солей и воды из почечных клеток, освобождение инсулина из панкреатических клеток, частоту сердцебиений.
Установлено, что энергетически выгодный транспорт ионов Na+ внутрь клеток оказывает также влияние на транспорт сахаров и аминокислот в клетки. В частности, с транспортом ионов Na+ сопряжен транспорт глюкозы. Чтобы создать градиент концентрации ионов Na+, благоприятный для транспорта ионов К+ и глюкозы внутрь клеток, ионная «насосная» система благодаря энергии активно откачивает ионы Na+ из клетки за ее пределы.
Определенная роль в транспорте веществ принадлежит белоксвязывающим системам, представляющим четвертый способ транспорта. Речь идет о белках, локализованных в периплазматическом пространстве. Эти белки специфически связывают сахара, аминокислоты и ионы, перенося их затем к специфическим молекулам-носителям, локализованным в клеточной мембране. Источником энергии для этих систем является АТФ.
Эндоцитоз, как отмечено выше, обеспечивает перенос в клетки крупных частиц и молекул. В рамках эндоцитоза различают фагоцитоз и пиноцитоз.
Фагоцитоз (от греч. phagos — пожирающий и cytos — клетка) представляет собой процесс, заключающийся в том, что клетки-лейкоциты (макрофаги и нейтрофилы) захватывают (обволакивают) твердые частицы (фрагменты клеток, бактерии) путем выпячиваний своей клеточной мембраны и образования пузырьков, сливающихся затем с плазматической мембраной и открывающихся внутрь клетки. Вошедшие внутрь клеток частицы поступают в лизосомы, где с помощью клеточных (лизосомных) ферментов разрушаются и усваиваются затем клетками. Фагоцитоз широко распространен среди одноклеточных организмов. У многоклеточных (млекопитающих) он выполняется специализированными клетками (лейкоцитами).
У простейших фагоцитоз является формой питания, в результате которого твердые частицы проникают в лизосомы, где и перевариваются, образуя продукты, служащие пищей. Биологическое значение фагоцитоза у млекопитающих заключается в том, что он обеспечивает иммунную (фагоцитарную) защиту организма (см. гл. XVII).
Пиноцитоз (от греч. pino — пить и cytos — клетка) представляет собой процесс, при котором клетки поглощают жидкости и находящиеся в них высокомолекулярные вещества путем впячива-ний плазматической мембраны и образования пузырьков (каналь-цев), куда поступает жидкость. Канальцы после заполнения жидкостью отшнуровываются, поступают в цитоплазму и доходят до лизосом, где их стенки перевариваются, в результате чего содержимое (жидкость) канальцев освобождается и подвергается дальнейшей обработке лизосомными ферментами.
Пиноцитоз часто встречается у одноклеточных животных, у многоклеточных он наблюдается в клетках кровеносной и лимфатической систем, в клетках злокачественных опухолей, а также в клетках тканей, для которых характерен повышенный уровень обмена веществ.
Экзоцитоз — это процесс секретирования клетками различных веществ, причем известны регулируемый и конститутивный пути экзоцитоза. Примером регулируемого экзоцитоза является экзоци-тоз инсулина. Клетки поджелудочной железы, продуцирующие инсулин, упаковывают его вначале в так называемые секреторные пузырьки, которые после внеклеточного сигнала сливаются с плазматической мембраной, а затем открываются в межклеточное пространство, освобождая гормон. Подобным образом происходит эк-зоцитоз других гормонов, нейротрансмиттеров и многих ферментов. Напротив, конститутивный путь экзоцитоза присущ многим белкам, непрерывно синтезируемым клетками и упаковываемым в эк-зоцитозные пузырьки в комплексе Гольджи, после чего эти пузырьки перемещаются к плазматической мембране, где и открываются в межклеточное пространство, освобождаясь от белкового содержимого.
С помощью экзоцитоза из клетки удаляются также частицы, оказавшиеся непереваренными путем фагоцитоза. У большинства клеток циклы эндоцитоз-экзоцитоз непрерывны.
Дата добавления: 2016-05-30; просмотров: 4834;
Поиск по сайту
Узнать еще
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I. Поверхностный аппарат клетки.
- I.2.2 ТЕПЛОЁМКОСТЬ ВЕЩЕСТВА
- VI. Пектиновые вещества и их роль в пищевой промышленности.
- А) Определение резистентности грудной клетки
- Аварийно химически опасные вещества (АХОВ)
- Агрегатные состояния вещества
- Активность радиоактивного вещества

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
